La storia dell'aspetto del soprannome
AGOSTO 2009 Chiudi finestra Ivan il Terribile, Pietro il Grande e Yaroslav il Saggio. Il suo soprannome è il Granduca di Kiev...
RIPARAZIONE DEL DNA
Sistemi di riparazione
2 Riparazione dell'escissione. Esempi e tipi
3 Riparazione degli errori di replicazione del DNA
4 Riparazione ricombinante (post-replicativa) nei batteri
5 Riparazione SOS
I sistemi di riparazione del DNA sono piuttosto conservativi nell'evoluzione dai batteri all'uomo e sono meglio studiati in E. coli.
Esistono due tipi di riparazione:
dritto ed escissionaleRiparazione diretta
La riparazione diretta è il modo più semplice per eliminare il danno nel DNA, che di solito coinvolge enzimi specifici che possono eliminare rapidamente (di solito in una fase) il danno corrispondente, ripristinando la struttura originale dei nucleotidi.
1. Questo funziona, ad esempio,O6-metilguanina-DNA-metiltransferasi
(un enzima suicida) che rimuove un gruppo metilico da una base azotata ad uno dei propri residui di cisteina
In E. coli, è possibile sintetizzare fino a 100 molecole di questa proteina in 1 minuto. La proteina degli eucarioti superiori, simile nella funzione, gioca ovviamente un ruolo importante nella protezione contro il cancro causato da fattori alchilanti interni ed esterni.
DNA inseriscesi
2. Insertasi di DNA
fotoliasi
3. I dimeri di timina sono "ricamati" da riparazione diretta protagonistafotoliasi, che effettuano la corrispondente trasformazione fotochimica. Le fotoliasi del DNA sono un gruppo di enzimi attivati dalla luce con una lunghezza d'onda di 300 - 600 nm (regione visibile), per i quali hanno uno speciale centro fotosensibile nella loro struttura.
Sono ampiamente distribuiti in natura e si trovano in batteri, lieviti, insetti, rettili, anfibi e esseri umani. Questi enzimi richiedono una varietà di cofattori (FADH, acido tetraidrofolico, ecc.) coinvolti nell'attivazione fotochimica dell'enzima. La fotoliasi di E. coli è una proteina di 35 kDa strettamente legata oligoribonucleotide lungo 10-15 nucleotidi necessaria per l'attività enzimatica.
Esempi di riparazione diretta
1. Base metilata O 6 mg dimetilato dall'enzima metiltransferasiO6-metilguanina-DNA-metiltransferasi (un enzima suicida) che trasferisce un gruppo metilico a uno dei suoi residui
cisteina
2. I siti AP possono essere riparati mediante l'inserimento diretto di purine con l'aiuto di enzimi chiamatiInsertasi di DNA(dall'inglese insert-insert).
SCHEMA DI UN ESEMPIO DI RIPARAZIONE DEL DANNO DIRETTO NEL DNA - Base metilata o6- mgdemetilato dall'enzima metiltransferasi, che trasferisce un gruppo metilico a uno dei suoi residui di amminoacido cisteina.
3. La fotoliasi si attacca al dimero di timina e, dopo l'irradiazione di questo complesso con luce visibile (300-600 nm), il dimero si espande
SCHEMA DI ESEMPIO DI RIPARAZIONE DIRETTA DI DANNI NEL DNA – Fotoliasi
si attacca al dimero di timina e, dopo irradiazione con luce visibile, questo dimero si espande

Riparazione dell'escissione
(dall'inglese escissione - taglio).
DEFINIZIONE
La riparazione dell'escissione include rimozione basi azotate danneggiate dal DNA e successivo recupero normale struttura molecolare.
MECCANISMO
Diversi enzimi sono solitamente coinvolti nella riparazione dell'escissione e il processo stesso influisce
non solo danneggiato ,
ma anche nucleotidi adiacenti .
CONDIZIONI
Per la riparazione dell'escissione, è necessario un secondo filamento (complementare) di DNA. Lo schema generale semplificato di riparazione escissionale è mostrato in fig. 171.
FASI
Il primo passo nella riparazione dell'escissione è l'escissione delle basi azotate anormali. È catalizzato da un gruppoDNA-N-glicosilasi- enzimi che scindono il legame glicosidico tra il desossiribosio e una base azotata.
NOTA IMPORTANTE:
InumanoDNA-N-glicosilasihanno un'elevata specificità di substrato: diversi enzimi di questa famiglia si riconoscono ed asportano varie basi anomale(8-ossoguanina, uracile, metilpurina, ecc.).
InbatteriDNA-N-glicosilasinon ha tale specificità del substrato.
ENZIMI GENERALI DI RIPARAZIONE DELL'ECCISIONE
| TITOLO | FUNZIONE | MECCANISMO |
| DNA-N-glicosilasi | escissione di basi azotate anomale | scinde il legame glicosidico tra il desossiribosio e base azotata |
| Endonucleasi AP | crea le condizioni per il lavoro del prossimo enzima - esonucleasi | rompe la spina dorsale zucchero-fosfato della molecola di DNA nel sito AP |
| esonucleasi | scinde alcuni nucleotidi | scinde in sequenza diversi nucleotidi dalla sezione danneggiata di un filamento di DNA |
PASSI SEQUENZIALI SPECIFICI DI QUESTA MECCANICA:
Come risultato dell'azione DNA- N-glicosilasisi forma un sito AP, che viene attaccato dall'enzima AR-endonucleasi. Rompe la spina dorsale zucchero-fosfato della molecola di DNA nel sito AP e quindi crea le condizioni per il lavoro del prossimo enzima - esonucleasi, che separa in sequenza diversi nucleotidi dalla sezione danneggiata di un filamento di DNA.
Nelle cellule batteriche lo spazio lasciato libero viene riempito con i nucleotidi corrispondenti con la partecipazione DNA polimerasi I orientato al secondo filamento (complementare) di DNA.
Poiché la DNA polimerasi I è in grado di estendere l'estremità 3' di uno dei filamenti in un'interruzione nel DNA a doppio filamento e rimuovere i nucleotidi dall'estremità 5' della stessa rottura,
quelli. rendersi conto “nick-trasmissione” , questo enzima svolge un ruolo chiave nella riparazione del DNA. Viene eseguita la cucitura finale delle aree riparate DNA ligasi.
Nelle cellule eucariotiche (mammiferi).
La riparazione dell'escissione del DNA nelle cellule di mammifero è accompagnata da un forte aumento dell'attività di un altro enzima,poli ADP-ribopolimerasi . Allo stesso tempo, succede ADP-ribosilazione delle proteine della cromatina(istoni e proteine non istoniche), che porta a un indebolimento della loro connessione con il DNA e apre l'accesso agli enzimi di riparazione.
Donatore ADP-ribosioin queste reazioniNAD+, le cui riserve sono notevolmente esaurite durante la riparazione escissionale dei danni causati dall'irradiazione di raggi X:

Residui caricati negativamente ADP-ribosio dalla composizione interna della molecola NAD+ aderire tramite un radicaleglutammina acidi o fosfoserinaalle proteine della cromatina, che porta alla neutralizzazione delle cariche positive di queste proteine e all'indebolimento del loro contatto con il DNA.
COS'È UN GRUPPO DI ENZIMI
DNA - glicosilasi
scinde il legame glicosidico tra desossiribosio e base azotata
che porta all'escissione di basi azotate anormali
DNA - glicosilasi , coinvolti nell'eliminazione del danno ossidativo al DNA nelle cellule procarioti ed eucarioti, sono molto diversi e differiscono per specificità del substrato, struttura spaziale e modalità di interazione con il DNA.
Le glicosilasi del DNA più studiate includono:
endonucleasi III(Endo III),
forma ammido pirimidina-DNA-glicosilasi (fpg)
Mu T e
Mu Ycoli.
Endonucleasi IIIE. coli riconosce e taglia in modo specifico dal DNA ossidato basi pirimidiniche.
Questo enzima è una proteina globulare monomerica costituita da 211 aminoacidi residui (peso molecolare 23,4 kDa). Il gene che codifica per Endo III è stato sequenziato ed è stata stabilita la sua sequenza nucleotidica. Endo III è proteina ferro-zolfo [(4 Fe-4S )2+-proteina], che contiene l'elemento struttura sovrasecondaria digitare "Chiave greca" (spirale - tornante - spirale), serve per legarsi al DNA. Sono stati isolati anche enzimi con una specificità di substrato simile e sequenze di amminoacidi simili cellule bovine e umane.
Forma ammido piridina-DNA-glicosilasi E. coli "riconosce" e scinde l'eterociclico ossidato basi della serie delle purine .
FASE DEL PROGRAMMA DI RIPARAZIONE DELL'ESCISIONE 1
DNAN

SCHEMA DI RIPARAZIONE DELL'ESCISIONE
1 DNANla glicosidasi rimuove la base danneggiata
L'endonucleasi AR rompe il DNA
2 L'esonucleasi rimuove un certo numero di nucleotidi
3 DNA polimerasi riempie il sito vuoto con complementare
Mononucleotidi
La DNA ligasi reticola il filamento di DNA riparato

Mu T- una piccola proteina con un peso molecolare di 15 kDa, che ha attività nucleosidica trifosfatasica, che è prevalentemente idrolizza dGTP in dGMP e pirofosfato.
Ruolo biologico del Mut T è impedire la formazione di coppie non canoniche durante la replicazioneA:G e R: 8-oxo-G.
Tali coppie possono apparire quando forma ossidata
dGTP (8-oxo-dGTP) diventa substrato DNA polimerasi.
Mu T idrolizza 8-oxo-dGTP10 volte più veloce di dGTP.
Lo fa 8-oxo-dGTPsubstrato più preferitoMutoTe ne spiega il ruolo funzionale.
Mu Yè una glicosilasi adenina-DNA specifica che scinde il legame N-glicosidico tra adenina e desossiribosio adenosina, formando una coppia non canonica con la guanina.
Il ruolo funzionale di questo enzima è prevenire la mutazione
T:A - G:A di taglio del residuo intatto adeninadalla coppia di basi A: 8-oxo-G.
Riparazione dell'escissione del nucleotide
(Meccanismo di rimozione del danno al DNA ATP-dipendente)
Ultimamente in riparazione per escissione Attenzione speciale forniscono un meccanismo ATP-dipendente per rimuovere il danno dal DNA. Questo tipo di riparazione per escissione è chiamata riparazione per escissione nucleotidica (NER).
Comprende DUE FASI :
1. rimozione dal DNAframmenti di oligonucleotidi contenente danni, e
Eccinucleasi
2. successiva ricostruzione della catena del DNA con la partecipazione di un complesso di enzimi (nucleasi, DNA polimerasi, DNA ligasi, ecc.).
Si verifica la rimozione di un frammento di DNA su entrambi i lati del danneggiato nucleotide. La lunghezza dei frammenti di oligonucleotidi rimossi differisce tra procarioti ed eucarioti.
Rimozione di un frammento di DNA dai procarioti
Quindi, in E. coli, B. subtilus, Micrococcus luteus, un frammento di lunghezza 12-13 nucleotidi
Rimozione di un frammento di DNA dagli eucarioti
e nel lievito, negli anfibi e nell'uomo, un frammento costituito da 24-32 nucleotidi.
Eccinucleasi- un enzima che rimuove i frammenti di DNA
Il frammento di DNA viene tagliato da un enzimaeccinucleasi(eccinucleasi). In E. coli, questo enzima è costituito da 3 diversi protomeri:
uvra
uvr B
uvr C
ognuno dei quali svolge una funzione specifica durante la scissione escissionale di un frammento di DNA. Il nome di queste proteine è dato dalle prime lettere delle parole"riparazione ultravioletta".
Protomer uvr Aha attività ATPasi, si lega al DNA sotto forma di un dimero, svolgendo
riconoscimento del danno primario e
rilegaturauvr B
Protomer uvr B ha:
uno . Latente Attività dell'ATPasi e dell'elicasi latente necessarie per modificare le conformazioni e svolgere la doppia elica del DNA;
2. Endonucleasi attività, scindendo il legame internucleotidico (fosfodiestere) dal latoZ"-fineframmento spaccato.
Protomer uvr Cagisce come endonucleasi, introducendo un'interruzione nel filamento di DNA riparato con5"-estremitàframmento tagliato.
Quindi i protomeriuvr A, uvr B, uvr Cinteragiscono con il DNA in una certa sequenza, effettuando una reazione ATP-dipendentescissione del frammento oligonucleotidico dal filamento di DNA che viene riparato.
Il divario risultante nella molecola del DNA viene riparato con la partecipazione della DNA polimerasi I e della DNA ligasi. Il modello di riparazione escissionale con la partecipazione degli enzimi di cui sopra è mostrato in fig. 172.
Riparazioni escissionali nell'uomo
Anche le riparazioni dell'escissione umana dipendono dall'ATP e includonotre fasi principali :
riconoscimento del danno,
doppio taglio del filamento di DNA,
sintesi riduttiva e
legatura del filo riparato.
Tuttavia, comporta la riparazione dell'escissione del DNA umano
25 diversi polipeptidi ,
16 di cui sono coinvolti nella scissione del frammento oligonucleotidico, essendo protomerieccinucleasi,
e il resto 9 effettuare la sintesi della porzione riparata della molecola.
Nel sistema di riparazione del DNA nell'uomo, un ruolo molto significativo è svolto dalle proteine di trascrizione -
RNA polimerasi II e
TF lununo dei sei principali fattori di trascrizione eucariota.
Va notato che la riparazione dell'escissione nei procarioti, così come negli eucarioti, dipende dallo stato funzionale del DNA:
il DNA trascritto viene riparato più velocemente
che trascrizionalmente inattivo.
Questo fenomeno è spiegato dai seguenti fattori:
struttura della cromatina,
omologia di catene di regioni di DNA trascritte,
effetto del danno alla catena e suo effetto sulla RNA polimerasi.
NOTA IMPORTANTE:
CODING CHAIN DNA (chain of information storage)
MATRIX CHAIN di DNA (le informazioni vengono cancellate da essa)
È noto che tali grandi danni come formazione del dimero di timina, bloccano la trascrizione sia nei batteri che nell'uomo se si verificano circuito a matrice DNA (danno a codifica Catene non intaccare al complesso di trascrizione). L'RNA polimerasi si ferma nel sito del danno al DNA e blocca il lavoro del complesso di trascrizione.
Fattore di collegamento trascrizione-riparazione (TRCF) .
In E. coli, il miglioramento della riparazione trascrizionale è mediato da una proteina speciale -fattore di collegamento trascrizione-riparazione (TRCF) .
Questa proteina contribuisce :
1. distacco dell'RNA polimerasi dal DNA
2. stimola contemporaneamente la formazione di un complesso proteico,
Effettuare la riparazione dell'area danneggiata.
Al termine della riparazione, l'RNA polimerasi si posiziona e la trascrizione continua (vedi Fig.).
Così schema generale riparazione dell'escissione
1. DNA-N -glicosilasi rimuove la base danneggiata
2. AR-endonucleasi introduce un'interruzione nella catena del DNA
3. L'esonucleasi rimuove un certo numero di nucleotidi
4. La DNA polimerasi riempie l'area libera
Nucleotidi complementari
5. La DNA ligasi lega in modo incrociato il filamento di DNA riparato
Riparazione di errori di replicazione del DNA
per metilazione
Errori nell'accoppiamento delle basi azotate durante la replicazione del DNA si verificano abbastanza spesso (nei batteri una volta ogni 10 mila nucleotidi), a seguito dei qualinel filamento figlia del DNA sono inclusi i nucleotidi che non sono complementari ai nucleotidi della catena materna -disallineamenti(Mismatch inglese n e partita).
Nonostante il fatto cheDNA polimerasi Iil procariota ha la capacità di autocorreggersi, i suoi sforzi per eliminare i nucleotidi attaccati erroneamente a volte non abbastanza, e poi alcune coppie sbagliate (non complementari) rimangono nel DNA.
In questo caso, la riparazione avviene utilizzando uno specifico sistema associatoMetilazione del DNA . L'azione di questo sistema di riparazione si basa sul fatto che dopo la replicazione, dopo un certo tempo (alcuni minuti), il DNA subisce la metilazione.
E.coli metilato in primis adenina con l'istruzione
N6-metil-adenina (N6-mA).
Fino a questo punto, il nuovo sintetizzato(filiale)la catena rimane non metilata.
Se ci sono nucleotidi spaiati in una tale catena, allora viene riparata: cosìla metilazione segna il DNA e
include un sistema di correzione degli errori replica.
In questo sistema riparativo si riconoscono strutture speciali:
sotto sequenzaG-N6-mA-TCe prossimo dietro di essa la deformazione
nella doppia elica nel sito di mancanza di complementarità (figura sotto).
nell'eliminazione dei nucleotidi spaiati in semi-metilato un complesso piuttosto complesso di enzimi di riparazione prende parte alla molecola di DNA, che scansiona la superficie della molecola di DNA,taglia una sezione della catena del bambino ricorrendo a disadattamentoe quindi crea le condizioni per la costruzione
i suoi nucleotidi desiderati (complementari).
Diversi componenti di questo complesso hanno attività diverse.nucleasi,
elicasi,
ATPaznoy,
necessario per introdurre rotture nel DNA e fendere i nucleotidi, svolgere la doppia elica del DNA e fornire energia per il movimento del complesso lungo la parte riparata della molecola.
Un complesso di enzimi di riparazione simili per struttura e funzioni è stato trovato anche nell'uomo.
Riparazione ricombinante (post-replicativa).
Nei casi in cui, per un motivo o per l'altro, i sistemi di riparazione di cui sopra vengono interrotti, possono formarsi delle lacune (aree non riparate) nelle catene del DNA, che hanno a volte abbastanza significativo, che è irto di interruzioni del sistema di replicazione e può portare alla morte cellulare.
In questo caso, la cellula è in grado di utilizzare per la riparazione di una molecola di DNA un'altra molecola di DNA ottenuta dopo la replicazione, cioè di coinvolgere a tal fine il meccanismori combinazione.
Nei batteri
Nei batteri interviene la riparazione ricombinanteRec Una proteina. Si lega a una regione di DNA a filamento singolo e lo coinvolge nella ricombinazione conregioni omologhe di filamenti intatti di un'altra molecola di DNA .
Di conseguenza, sia le catene rotte (contenenti spazi vuoti) che intatte della molecola di DNA riparata risultano essere accoppiato con regioni di DNA complementari intatte, che apre la possibilità di riparazione mediante i sistemi sopra descritti.
Allo stesso tempo, potrebbe esserci tagliare frammento specifico e
Riempimentocon il suo aiuto lacune nel circuito difettoso.
Le lacune e le rotture risultanti nelle catene del DNA sono colmate con la partecipazione diDNA polimerasi I e DNA ligasi .
SOS riparazione
L'esistenza di questo sistema è stata postulata per la prima volta da M. Radman nel 1974. Ha anche dato il nome a questo meccanismo includendo in esso il segnale di soccorso internazionale "SOS" (salva le nostre anime).
In effetti, questo sistema si accende quando il danno al DNA diventa così grande da minacciare la vita della cellula. In questo caso, viene indotta l'attività di un gruppo eterogeneo di geni coinvolti in vari processi cellulari associati alla riparazione del DNA.
L'inclusione di determinati geni, determinata dalla quantità di danno nel DNA, porta a risposte cellulari di diverso significato (a partire dallo standard riparazione del danneggiato nucleotidi e desinenza soppressione divisione cellulare).
I più studiatiSOS riparazionein E. coli, i cui partecipanti principali sono proteine codificate geni Rec AeLex A.
Il primo è un multifunzionaleRec Una proteina partecipando
in Ricombinazione del DNA, così come
in regolazione della trascrizione genica fago lambdache infetta E. coli,
e secondo (proteina Lex A)è un repressore trascrizione di un grande gruppo di geni destinati Riparazione del DNA batteri. Quando è inibito o risolto la riparazione è attivata.
Rilegatura Rec A con Lex Aconduce alla scissione di questi ultimi e di conseguenza a attivazione dei geni di riparazione.
A sua volta, serve l'induzione del sistema SOS battericofago lambda segnale di pericolo e fa passare il profago da percorso da passivo ad attivo (litico). esistenza, causando così morte della cellula ospite.
Il sistema di riparazione SOS è stato trovato non solo nei batteri, ma anche negli animali e nell'uomo.
Geni coinvolti nella riparazione del danno del DNA SOS
| Geni | Conseguenze dell'attivazione genica |
| uvr A, B, C, D | Riparazione del danno alla struttura secondaria del DNA |
| Rec A | Riparazione post-replicativa, induzioni del sistema SOS |
| lex A | Spegnimento del sistema SOS |
| rec N, ruv | Riparazione di rotture a doppio filamento |
| Garantire la riparazione della ricombinazione |
|
| umu C, D | Mutagenesi causata da cambiamenti nelle proprietà della DNA polimerasi |
| sul A | Soppressione della divisione cellulare |
Conclusione
La riparazione del danno al DNA è strettamente correlata ad altri processi genetici molecolari fondamentali: replicazione, trascrizione e ricombinazione. Tutti questi processi lo sono intrecciate in un sistema comune di interazioni servito da un gran numero di proteine diverse, molte delle quali sono molecole polifunzionali coinvolte controllo sull'implementazione dell'informazione genetica nelle cellule pro ed eucariotiche. Allo stesso tempo, è chiaro che la natura "non lesinare" sugli elementi di controllo, creando i sistemi più complessi per correggere quei danni nel DNA che sono pericolosi per l'organismo e soprattutto per la sua progenie. D'altra parte, nei casi in cui la capacità riparativa non è sufficiente a mantenere lo stato genetico dell'organismo, è necessaria la morte cellulare programmata -apoptosi..
SCHEMA DI RIPARAZIONE DELL'ECCISIONE DEI NUCLEOTIDI IN e. COLCON LA PARTECIPAZIONE DI EXINUCLEASE
1. MECCANISMO INDIPENDENTE DALLA TRASCRIZIONE

2. MECCANISMO DIPENDENTE DALLA TRASCRIZIONE

3. FASE GENERALE DI RIPARAZIONE

SIMBOLI
A - proteineuvr MA
B - proteineuvr A
C - proteinauvr Insieme a
piccolo triangolo nero: il segno indica la posizione del danno
SCHEMA DI RIPARAZIONE ASSOCIATO ALLA METILAZIONE DEL DNA

accoppia gli altri. Questi cambiamenti accompagnano ogni ciclo di replicazione del DNA, ma la loro frequenza è molto inferiore a quella che dovrebbe essere. Ciò è spiegato dal fatto che la maggior parte dei cambiamenti di questo tipo vengono eliminati a causa dell'azione del meccanismo di riparazione (ripristino molecolare) della sequenza nucleotidica del DNA originale.
Il meccanismo di riparazione si basa sulla presenza di due catene complementari nella molecola del DNA. La distorsione della sequenza nucleotidica in uno di essi viene rilevata da enzimi specifici. Quindi il sito corrispondente viene rimosso e sostituito da uno nuovo, sintetizzato sul secondo filamento di DNA complementare. Tale riparazione è chiamata escissionale, cioè con "taglio" (Fig. 3.15). Viene eseguito prima del successivo ciclo di replica, quindi viene anche chiamato
pre-replicativo.
Riso. 3.14. Schema del processo di correzione durante la sintesi del DNA: I - inclusione nella catena del DNA di un nucleotide con una forma modificata (tautomerica) di citoeina, che si accoppia "illegalmente" con l'adenina; II - transizione veloce
la citosina nella sua forma abituale interrompe la sua associazione con l'adenina; l'estremità 3"-OH spaiata della catena sintetizzata ne impedisce l'ulteriore allungamento sotto l'azione della DNA polimerasi; III - la DNA polimerasi rimuove il nucleotide illegale, a seguito del quale riappare l'estremità 3"-OH accoppiata con lo stampo; IV - La DNA polimerasi continua a costruire la catena all'estremità 3 "-OH
Il ripristino della struttura originale del DNA richiede la partecipazione di numerosi enzimi. Un punto importante nell'avvio del meccanismo di riparazione è il rilevamento di un errore nella struttura del DNA. Spesso tali errori si verificano nel filamento appena sintetizzato durante la replicazione. Gli enzimi di riparazione devono rilevare esattamente questa catena. In molte specie di organismi viventi, la catena del DNA di nuova sintesi differisce dal grado materno di metilazione delle sue basi azotate, che è in ritardo rispetto alla sintesi. In questo caso, la catena non metilata viene riparata. L'oggetto del riconoscimento da parte degli enzimi di riparazione può anche essere l'interruzione della catena del DNA. Negli organismi superiori, dove la sintesi del DNA non avviene continuamente, ma da singoli repliconi, la catena del DNA appena sintetizzata presenta rotture, che ne consentono il riconoscimento.
Il ripristino della struttura del DNA in caso di perdita delle basi puriniche di una delle sue catene comporta l'individuazione di un difetto mediante l'enzima endonucleasi, che rompe il legame fosfoestere nel sito di danneggiamento della catena. Quindi il sito alterato con diversi nucleotidi adiacenti viene rimosso dall'enzima esonucleasi e al suo posto, secondo l'ordine delle basi della catena complementare, si forma la sequenza nucleotidica corretta (Fig. 3.15).

Riso. 3.15. Schema di riparazione del DNA escissionale e pre-replicativa Quando una delle basi nella catena del DNA viene modificata nel ripristino dell'originale
alle strutture partecipano circa 20 enzimi della DNA glicosilasi che riconoscono in modo specifico i danni causati dalla deaminazione, dall'alchilazione e da altre trasformazioni strutturali delle basi. Tali basi modificate vengono rimosse. Ci sono aree prive di basi, che vengono riparate, come per la perdita di purine. Se non viene effettuato il ripristino della struttura normale, ad esempio, nel caso di deaminazione di basi azotate, alcune coppie di basi complementari vengono sostituite da altre - la coppia C-G può essere sostituita coppia T-A eccetera. (Vedere la sezione 3.4.2.3).
La formazione di dimeri di timina (T-T) in catene polinucleotidiche sotto l'azione dei raggi UV richiede la partecipazione di enzimi che riconoscono non singole basi alterate, ma danni più estesi alla struttura del DNA. Il processo riparativo in questo caso è anche associato alla rimozione del sito che porta il dimero e al ripristino della normale sequenza nucleotidica per sintesi sul filamento di DNA complementare.
Nel caso in cui il sistema di riparazione dell'escissione non corregga un cambiamento che si è verificato in un filamento di DNA, la fissazione si verifica durante la replicazione.
questo cambiamento e diventa proprietà di entrambi i filamenti di DNA. Ciò porta alla sostituzione di una coppia di nucleotidi complementari con un'altra o alla comparsa di rotture (lacune) nella catena di nuova sintesi contro le regioni alterate. Il ripristino della normale struttura del DNA può avvenire anche dopo la replicazione.
Riparazione post-replicativa avviene per ricombinazione (scambio di frammenti) tra due doppie eliche di DNA di nuova formazione. Un esempio di tale riparazione post-replicativa è il ripristino della normale struttura del DNA quando compaiono i dimeri di timina (T-T), quando non vengono eliminati spontaneamente sotto l'azione della luce visibile ( riparazione leggera) o durante la riparazione dell'escissione pre-replicativa.
I legami covalenti che si verificano tra i residui di timina adiacenti li rendono incapaci di legarsi ai nucleotidi complementari. Di conseguenza, nel filamento di DNA appena sintetizzato compaiono rotture (lacune) riconosciute dagli enzimi di riparazione. Il ripristino dell'integrità della nuova catena polinucleotidica di uno dei DNA figli viene effettuato a causa della ricombinazione con la corrispondente catena materna normale dell'altro DNA figlia. Il gap formatosi nella catena madre viene quindi riempito mediante sintesi sulla catena polinucleotidica complementare (Fig. 3.16). Una manifestazione di tale riparazione post-replicazione, effettuata mediante ricombinazione tra le catene di due molecole di DNA figlie, può essere considerata lo scambio di materiale spesso osservato tra cromatidi fratelli (Fig. 3.17).

Riso. 3.16. Schema di riparazione del DNA postreplicativo: I - presenza di dimero di timina in uno dei filamenti di DNA;
II - la formazione di un "gap" nel filamento appena sintetizzato contro il sito alterato della molecola madre dopo la replicazione (la freccia mostra il successivo riempimento del "gap" con un sito dal filamento corrispondente della seconda molecola di DNA figlia) ;
III - ripristino dell'integrità della catena figlia della molecola superiore per ricombinazione e nella molecola inferiore per sintesi sulla catena complementare

Riso. 3.17. Scambi intercromatidi (indicati dalle frecce)
Nel corso della riparazione pre-replicativa e post-replicativa, la maggior parte del danno alla struttura del DNA viene ripristinato. Tuttavia, se si verificano troppi danni nel materiale ereditario della cellula e alcuni di essi non vengono eliminati, il sistema degli enzimi di riparazione inducibili (eccitati) (sistema SOS) viene attivato. Questi enzimi colmano le lacune ripristinando l'integrità delle catene polinucleotidiche sintetizzate senza osservare rigorosamente il principio di complementarità. Ecco perché a volte i processi di riparazione stessi possono fungere da fonte di cambiamenti persistenti nella struttura del DNA (mutazioni). La reazione denominata si applica anche al sistema SOS.
Se nella cellula, nonostante la riparazione in corso, la quantità di danno alla struttura del DNA rimane elevata, i processi di replicazione del DNA vengono bloccati in essa. Tale cellula non si divide, il che significa che non trasmette i cambiamenti che sono sorti alla prole.
L'arresto del ciclo cellulare causato dal danno del DNA, unito all'impossibilità di riparazione molecolare del materiale ereditario alterato, può, con l'intervento di una proteina la cui sintesi è controllata dal gene p53, portare all'attivazione del processo di autodeterminazione -distruzione (apoptosi) di una cellula difettosa per eliminarla dall'organismo.
Pertanto, un vasto insieme di vari enzimi di riparazione esegue un continuo "esame" del DNA, rimuovendo le aree danneggiate da esso e contribuendo a mantenere la stabilità del materiale ereditario. L'azione congiunta degli enzimi di replicazione (DNA polimerasi ed endonucleasi di editing) e degli enzimi di riparazione assicura un tasso di errore abbastanza basso nelle molecole di DNA, che viene mantenuto al livello di 1 × 10–9 coppie di nucleotidi alterati per genoma. Con una dimensione del genoma umano di 3 × 109 paia di basi, ciò significa la comparsa di circa 3 errori per replicazione del genoma. Allo stesso tempo, anche questo livello è sufficiente per la formazione di una significativa diversità genetica sotto forma di mutazioni genetiche durante l'esistenza della vita sulla Terra.

3.4.2.3. Cambiamenti nelle sequenze nucleotidiche del DNA. Mutazioni geniche
I cambiamenti non corretti nella struttura chimica dei geni, riprodotti in cicli successivi di replicazione e manifestati nella prole sotto forma di nuove varianti di tratti, sono chiamati mutazioni genetiche.
I cambiamenti nella struttura del DNA che compongono un gene possono essere divisi in tre gruppi. Le mutazioni del primo gruppo consistono nella sostituzione di alcune basi con altre. Costituiscono circa il 20% dei cambiamenti genetici spontanei. Il secondo gruppo di mutazioni è causato da uno spostamento del frame che si verifica quando il numero di coppie di nucleotidi nel gene viene modificato. Infine, il terzo gruppo è rappresentato dalle mutazioni associate a un cambiamento nell'ordine delle sequenze nucleotidiche all'interno di un gene (inversione).
Mutazioni a seconda del tipo di sostituzione delle basi azotate. Queste mutazioni si verificano per una serie di ragioni specifiche. Uno di questi può essere un cambiamento nella struttura di una base che è già inclusa nell'elica del DNA, che si verifica per caso o sotto l'influenza di specifici agenti chimici. Se una tale forma alterata della base rimane inosservata dagli enzimi di riparazione, durante il successivo ciclo di replicazione può attaccare a se stessa un altro nucleotide. Un esempio è la deaminazione della citosina, che si trasforma in uracile spontaneamente o sotto l'influenza dell'acido nitroso (Fig. 3.18). L'uracile risultante, non notato dall'enzimaDNA glicosilasi,durante la replicazione, si combina con l'adenina, che successivamente si lega a un nucleotide timidilico. Di conseguenza, una coppia C-G è sostituito nel DNA da una coppia T-A (Fig. 3.19, I ). La deaminazione della citosina metilata la converte in timina (vedi Figura 3.18). Il nucleotide timidilico, essendo un componente naturale del DNA, non viene rilevato come un cambiamento dagli enzimi di riparazione e aggiunge un nucleotide adenile durante la successiva replicazione. Di conseguenza, invece di una coppia C-G una coppia appare anche nella molecola del DNA TA (Fig. 3.19, II).
Riso. 3.18. Deaminazione spontanea della citosina
Un'altra ragione per la sostituzione delle basi potrebbe essere l'errata inclusione nella catena del DNA sintetizzato di un nucleotide che porta una forma chimicamente modificata della base o un suo analogo. Se questo errore non viene notato dagli enzimi di replicazione e riparazione, la base modificata viene inclusa nel processo di replicazione, che spesso porta alla sostituzione di una coppia con un'altra. Un esempio di ciò è l'aggiunta di un nucleotide con 5-bromouracile (5-BU), simile al nucleotide timidilico, all'adenina della catena materna durante la replicazione. Durante la successiva replicazione, la 5-BU si attacca più facilmente non adenina, ma guanina. La guanina nel corso di un ulteriore raddoppio forma una coppia complementare con la citosina. Di conseguenza, la coppia AT viene sostituita nella molecola di DNA dalla coppia G-C (Fig. 3.20).

Riso. 3. 19. Mutazioni per tipo di sostituzione delle basi (deaminazione delle basi azotate nella catena del DNA):
I - conversione della citosina in uracile, sostituzione della coppia C-G con una coppia T-A;
II - conversione della metilcitosina in timina, sostituzione della coppia CG con una coppia TA
Dagli esempi di cui sopra, si può vedere che cambiamenti nella struttura della molecola di DNA in base al tipo di sostituzione delle basi si verificano prima o durante la replicazione, inizialmente in una catena polinucleotidica. Se tali modifiche non vengono corrette durante la riparazione, durante la successiva replicazione diventano proprietà di entrambi i filamenti di DNA.
Riso. 3.20. Mutazioni per sostituzione di basi
(Inclusione di un analogo di una base azotata nella replicazione del DNA)
La conseguenza della sostituzione di una coppia di nucleotidi complementari con un'altra è la formazione di una nuova tripletta nella sequenza nucleotidica del DNA che codifica la sequenza di amminoacidi nella catena peptidica. Ciò potrebbe non influire sulla struttura del peptide se la nuova tripletta è un "sinonimo" della precedente, cioè codificherà per lo stesso amminoacido. Ad esempio, l'amminoacido valina è crittografato con quattro triplette: CAA, CAG, CAT, CAC. Sostituire la terza base in una qualsiasi di queste triplette non ne cambierà il significato (la degenerazione del codice genetico).
A nel caso in cui la tripletta appena emersa codifichi un altro amminoacido, la struttura della catena peptidica e le proprietà della corrispondente proteina cambiano. A seconda della natura e del luogo della sostituzione, le proprietà specifiche della proteina cambiano in varia misura. Sono noti casi in cui la sostituzione di un solo amminoacido in un peptide influisce in modo significativo sulle proprietà della proteina, che si manifesta in un cambiamento di caratteristiche più complesse. Un esempio è il cambiamento nelle proprietà dell'emoglobina umana quando anemia falciforme (Fig. 3.21). In tale emoglobina-(HbS) (a differenza della normale HbA) - nelle catene p-globiniche nella sesta posizione, l'acido glutammico viene sostituito dalla valina. Questa è una conseguenza della sostituzione di una delle basi nella tripletta che codifica per l'acido glutammico (CTT o CTC). Di conseguenza, viene visualizzata una tripletta di crittografia valina (CAT o CAC). In questo caso, la sostituzione di un amminoacido nel peptide cambia significativamente le proprietà della globina, che fa parte dell'emoglobina (la sua capacità di legarsi a 02 diminuisce), una persona sviluppa segni di anemia falciforme.
A in alcuni casi, la sostituzione di una base con un'altra può portare alla comparsa di una di triplette senza senso (ATT, ATC, ACT) che non codificano per alcun amminoacido. La conseguenza di tale sostituzione sarà l'interruzione della sintesi della catena peptidica. Si stima che le sostituzioni nucleotidiche in una tripletta portino nel 25% dei casi alla formazione di triplette sinonime; in 2-3 triplette senza significato, nel 70-75% - al verificarsi di vere mutazioni genetiche.
Pertanto, le mutazioni di sostituzione delle basi possono verificarsi sia come risultato di cambiamenti spontanei nella struttura di base in uno dei filamenti di una doppia elica di DNA già esistente, sia durante la replicazione in un filamento di nuova sintesi. Se queste modifiche non vengono corrette durante la riparazione (o, al contrario, si verificano durante la riparazione), vengono fissate in entrambe le catene e verranno quindi riprodotte nei successivi cicli di replica. Pertanto, un'importante fonte di tali mutazioni sono le violazioni dei processi di replicazione e riparazione.
Mutazioni con uno spostamento nel frame di lettura. Questo tipo di mutazione costituisce una percentuale significativa di mutazioni spontanee. Si verificano a causa della perdita o dell'inserimento di una o più coppie di nucleotidi complementari nella sequenza nucleotidica del DNA. La maggior parte delle mutazioni studiate che causano
Indice del soggetto "Elementi genetici dei batteri. Mutazioni nei batteri. Trasduzione.":Ci sono meccanismi nella cellula che possono ripristinare completamente o parzialmente la struttura originale del DNA alterato. Mutazioni causate da radiazioni, sostanze chimiche e altri fattori potrebbero teoricamente portare all'estinzione della popolazione batterica se quest'ultima fosse privata della capacità di Riparazione del DNA. L'insieme degli enzimi che catalizzano la correzione del danno al DNA è combinato nei cosiddetti sistemi di riparazione, che differiscono fondamentalmente nei meccanismi biochimici di "guarigione" del danno. Ci sono tre direzioni principali per la correzione dei difetti del DNA.
1. Reversione diretta da DNA danneggiato alla struttura originaria quando cambiamenti nel DNA corretto da una singola reazione enzimatica. Ad esempio, rimozione di un gruppo metilico attaccato in modo errato al sesto atomo di ossigeno della guanina usando la metiltransferasi; o scissione del dimero di timina irradiato da fotoliasi ( riparazione di ricombinazione).
2. Danno da "taglio". seguito da ripristino della struttura originaria (riparazione mediante escissione).
3. Attivazione di meccanismi speciali che garantiscono la sopravvivenza in caso di danno DNA(restauro della struttura originaria DNA come risultato della ricombinazione; correzione della mancata corrispondenza di base; sintesi traslazionale su matrice danneggiata DNA). Questi meccanismi non portano sempre al completo ripristino della struttura originale del DNA.
mutazione primaria può essere compensato da una mutazione secondaria che si è verificata all'interno del gene mutato (per via intragena) o in un altro gene (per via extragena). Cambiamenti che eliminano le manifestazioni di una mutazione senza correggere il disturbo originario in DNA, chiamata soppressione.
soppressione intragenica causato da una mutazione secondaria che corregge gli effetti della mutazione primaria. Ad esempio, una mutazione puntiforme risultante nella sintesi di una proteina difettosa con attività biologica persa può essere corretta se una mutazione puntiforme secondaria risulta nella codificazione di un amminoacido che mantiene la configurazione e l'attività della proteina. Un esatto ripristino della struttura originale di un gene è chiamato una vera mutazione posteriore ( vero ritorno). Se l'effetto della prima mutazione è compensato da una mutazione in un'altra parte del gene, tali mutazioni sono chiamate reversioni secondarie.
Soppressione extragenica- soppressione della manifestazione di una mutazione avvenuta in un gene, a causa di una mutazione nel secondo gene.
Il meccanismo di riparazione si basa sulla presenza di due catene complementari nella molecola del DNA. La distorsione della sequenza nucleotidica in uno di essi viene rilevata da enzimi specifici. Quindi il sito corrispondente viene rimosso e sostituito da uno nuovo, sintetizzato sul secondo filamento di DNA complementare. Tale riparazione è chiamata escissionale, cioè con "taglio" (Fig. 15). Viene eseguito prima del successivo ciclo di replica, quindi è anche chiamato pre-replicativo.
Fig.14. Schema del processo di correzione durante la sintesi del DNA:
I-inclusione nella catena del DNA di un nucleotide con una forma modificata (tautomerica) di citoeina, che si accoppia "illegalmente" con l'adenina; II - la rapida transizione della citosina alla sua forma normale interrompe il suo accoppiamento con l'adenina; l'estremità 3"-OH spaiata della catena sintetizzata ne impedisce l'ulteriore allungamento sotto l'azione della DNA polimerasi; III - la DNA polimerasi rimuove il nucleotide illegale, a seguito del quale riappare l'estremità 3"-OH accoppiata con lo stampo; IV - La DNA polimerasi continua ad estendere la catena all'estremità 3'-OH.
Il ripristino della struttura originale del DNA richiede la partecipazione di numerosi enzimi. Un punto importante nell'avvio del meccanismo di riparazione è il rilevamento di un errore nella struttura del DNA. Spesso tali errori si verificano nel filamento appena sintetizzato durante la replicazione. Gli enzimi di riparazione devono rilevare esattamente questa catena. In molte specie di organismi viventi, la catena del DNA di nuova sintesi differisce dal grado materno di metilazione delle sue basi azotate, che è in ritardo rispetto alla sintesi. In questo caso, la catena non metilata viene riparata. L'oggetto del riconoscimento da parte degli enzimi di riparazione può anche essere l'interruzione della catena del DNA. Negli organismi superiori, dove la sintesi del DNA non avviene continuamente, ma da singoli repliconi, la catena del DNA appena sintetizzata presenta rotture, che ne consentono il riconoscimento. Il ripristino della struttura del DNA in caso di perdita delle basi puriniche di una delle sue catene comporta l'individuazione di un difetto mediante l'enzima endonucleasi, che rompe il legame fosfoestere nel sito di danneggiamento della catena. Quindi il sito alterato con diversi nucleotidi adiacenti viene rimosso dall'enzima esonucleasi e al suo posto, secondo l'ordine delle basi della catena complementare, si forma la sequenza nucleotidica corretta (Fig. 15).

Fig.15. Schema di riparazione del DNA escissionale e pre-replicativa.
Quando una delle basi della catena del DNA cambia, circa 20 enzimi della DNA glicosilasi partecipano al ripristino della struttura originaria e riconoscono in modo specifico i danni causati dalla deaminazione, dall'alchilazione e da altre trasformazioni strutturali delle basi. Tali basi modificate vengono rimosse. Ci sono aree prive di basi, che vengono riparate, come per la perdita di purine. Se il ripristino della struttura normale non viene eseguito, ad esempio, nel caso di deaminazione di basi azotate, alcune coppie di basi complementari vengono sostituite da altre - la coppia C-G può essere sostituita da una coppia T-A, ecc. .
La formazione di dimeri di timina (T-T) in catene polinucleotidiche sotto l'azione dei raggi UV richiede la partecipazione di enzimi che riconoscono non singole basi alterate, ma danni più estesi alla struttura del DNA. Il processo riparativo in questo caso è anche associato alla rimozione del sito che porta il dimero e al ripristino della normale sequenza nucleotidica per sintesi sul filamento di DNA complementare.
Nel caso in cui il sistema di riparazione dell'escissione non corregga un cambiamento che si è verificato in un filamento di DNA, durante la replicazione questo cambiamento viene fissato e diventa proprietà di entrambi i filamenti di DNA. Ciò porta alla sostituzione di una coppia di nucleotidi complementari con un'altra o alla comparsa di rotture (lacune) nella catena di nuova sintesi contro le regioni alterate. Il ripristino della normale struttura del DNA può avvenire anche dopo la replicazione.
Sotto l'azione di agenti alchilanti, ad esempio, nel DNA si formano residui di guanina N-metil-rTG-nitrosourea o H^gH-dimetilnitrosoguanidina, O 6 -metil - o O 6 -alchil-sostituiti. Tali residui di guanina modificati possono essere dealchilati da enzimi presenti nelle cellule batteriche e di mammifero. О 6 -metilguanina-DNA-alchiltransferasi catalizza il trasferimento di gruppi alchilici a gruppi sulfidrilici di cisteina residui dell'enzima, mentre la proteina accettore è inattivata. Il contenuto di alchiltransferasi nelle cellule di E. coli aumenta in presenza di O 6 -alchilguanina, ma non si osserva una simile inducibilità nelle cellule di mammifero.
Quando il DNA viene irradiato con luce ultravioletta, al suo interno si formano dimeri di ciclobutano tra basi pirimidiniche adiacenti. Tali composti bloccano la replicazione del DNA e devono essere rimossi per preservare la vitalità cellulare. Un modo per rimuovere i dimeri della pirimidina è convertirli enzimaticamente in monomeri illuminando la soluzione con luce visibile nell'intervallo di lunghezze d'onda di 300-600 nm. Tali enzimi fotoriattivanti si trovano nei batteri e negli organismi eucariotici inferiori, ma non sono stati trovati nelle cellule dei mammiferi. L'enzima forma un complesso stabile con il dimero pirimidinico e, utilizzando l'energia della luce da esso assorbita, distrugge il dimero senza rompere le catene del DNA.
La sostituzione di un nucleotide modificato di solito avviene in quattro fasi. In primo luogo, l'enzima riconosce questo nucleotide e taglia la catena polinucleotidica vicino ad esso o rompe il legame glicosidico tra la base modificata e il desossiribosio. In secondo luogo, l'esonucleasi rimuove il nucleotide modificato e/o i nucleotidi adiacenti, lasciando un piccolo spazio vuoto. In terzo luogo, il sito cancellato viene risintesi dal terminale 3-OH utilizzando il filamento opposto come modello. In quarto luogo, le estremità di interruzione risultanti dalla riparazione sono collegate per ripristinare l'integrità covalente del filo riparato.
I siti in cui si è verificata la depurinazione o la depirimidinazione vengono eliminati da enzimi chiamati endonucleasi AP. Nelle cellule pro ed eucariotiche, ci sono molte diverse endonucleasi AP, spesso diverse tipi diversi. Alcune endonucleasi AP si incidono dal lato 3' del sito AP, mentre altre scindono il legame diestere dal lato 3'; in entrambi i casi si formano le estremità 3'-idrossile e 5'-fosforile. La rottura del legame fosfodiestere su entrambi i lati del sito dell'interruzione consente all'esonucleasi di rimuovere i residui adiacenti su entrambi i lati del sito di scissione e quindi sintetizzare nuovamente la sequenza deleta.
Nella riparazione delle purine N-alchilate e di altre basi modificate, un ruolo chiave è svolto dalle N-glicosilasi specifiche, enzimi che scindono il legame glicosidico tra basi modificate e desossiribosio. Le N-glicosilasi trovano impiego anche nella correzione di disturbi consistenti nella deaminazione spontanea della citosina o dell'adenina e nella loro trasformazione, rispettivamente, in uracile o in ipoxantina. Gli enzimi uracile glicosilasi e ipoxantina glicosilasi scindono rispettivamente l'uracile e l'ipoxantina dal DNA, lasciando spazi vuoti nel sito di scissione. E ancora, utilizzando il meccanismo della scissione-risintesi, viene ripristinata la sequenza originale della catena danneggiata.
L'uracile glicosilasi svolge un ruolo antimutageno molto importante nella correzione degli errori che si verificano quando si utilizza dUTP invece di dTTP durante la replicazione. Poiché l'accoppiamento dUMP con il modello è buono quasi quanto dTMP, Pol III non lo riconosce e se dUMP non viene specificamente scisso, dGMP può essere incluso nel nuovo filamento durante un successivo round di replica. Pertanto, la g-glicosilasi protegge le cellule dalle possibili conseguenze mutagene dell'incorporazione di dUMP nella catena del DNA.
La riparazione dei dimeri di timina può avvenire anche al buio in uno dei due modi seguenti. Nelle cellule di E. coli vengono sintetizzati tre polipeptidi, codificati dai geni uvrA, uvrB e uvrC, che formano il complesso enzimatico uvrLVC-endonucleasi. Questo enzima taglia un filamento di DNA contenente un dimero pirimidinico a una distanza di otto legami fosfodiestere sul lato 5' e quattro o cinque legami sul lato 3' del dimero pirimidinico. La rimozione del sito danneggiato sembra essere catalizzata da un'elicasi codificata da un altro gene, uvrD. Questo rimuove il dimero di timina e circa altri 12 nucleotidi che lo circondano. Il divario risultante viene riempito con Pol I e la DNA ligasi completa la riparazione collegando due basi adiacenti della catena. Alcuni organismi utilizzano un meccanismo diverso per riparare i danni derivanti dalla formazione di dimeri di pirimidina. La riparazione viene eseguita con la partecipazione della pirimidina dimer-g-glicosilasi, che crea un sito apirimidinico. Il legame glicosidico tra una delle timine e il desossiribosio viene tagliato in modo che il dimero di timina rimanga all'estremità 5' della catena spezzata e il gruppo 3'-OH rimanga al desossiribosio. L'estremità 3'-AP risultante viene tagliata via dall'attività 3'-5'-esonucleasi della DNA polimerasi, quindi il nucleotide viene rimosso insieme all'adiacente dimero di timina mediante traslazione e legatura, lo spazio risultante viene riempito e le estremità della catena vengono suturate.
Nel processo di evoluzione, le cellule hanno sviluppato un meccanismo complesso per riparare i danni che si verificano nel DNA sotto l'influenza di un'ampia varietà di fattori chimici e fisici, nonché a causa di errori di replicazione o ricombinazione. E questo è abbastanza comprensibile: la maggior parte dei danni blocca il trasferimento di informazioni genetiche alla generazione successiva e il resto, se non viene eliminato, rimarrà nei genomi dei discendenti e porterà a cambiamenti drammatici nelle molecole proteiche, inclusi gli enzimi necessario per mantenere la vita della cellula. Quando alcune parti del sistema di riparazione vengono danneggiate, le cellule diventano particolarmente vulnerabili a determinati agenti chimici e fisici. Ad esempio, le cellule di E. coli, in cui il sistema per introdurre rotture nel DNA durante la scissione dei dimeri di timina, è molto sensibile alla luce UV. Le cellule che non sono in grado di eseguire questa o quella reazione della N-glicosilasi sono molto più suscettibili delle cellule normali all'effetto mutageno o letale degli agenti alchilanti o delle radiazioni ionizzanti. Nelle cellule di E. coli difettose in Pol I, la sopravvivenza è significativamente ridotta se irradiate con basse dosi di luce UV.
L'eucariotico inferiore Saccharomyces cerevisiae ha almeno cinque geni che codificano per proteine coinvolte nell'introduzione di rotture nel DNA irradiato dai raggi UV. Un'interruzione in uno solo di questi cinque geni RAD fa sì che le cellule perdano la capacità di introdurre rotture del DNA e quindi rimuovere i dimeri della pirimidina. Nel lievito, ci sono anche mutanti con una ridotta capacità di rimuovere i legami incrociati tra le catene, sebbene l'eliminazione del danno indotto dai raggi UV avvenga normalmente. Ciò suggerisce che nel lievito, come nell'uomo, esistono meccanismi di riparazione specifici e altamente complessi per rimuovere i legami incrociati e possibilmente per correggere molte altre modificazioni chimiche nel DNA.
Le persone con xeroderma pigmentoso sono molto sensibili alla luce ultravioletta e si sviluppano forme diverse cancro della pelle anche con pochissima esposizione alla luce solare. Le cellule di questi individui portano una mutazione simile alla mutazione del lievito RAD, in quanto hanno una ridotta capacità di scindere i dimeri della pirimidina dal DNA irradiato dai raggi UV. La malattia potrebbe essere dovuta a una mutazione in uno di almeno nove geni, il che indica un meccanismo piuttosto complesso per la riparazione del DNA contenente dimeri di timina nell'uomo. Di norma, la malattia è associata all'incapacità di scindere i dimeri di timina. Se un enzima con attività di timina dimer glicosilasi ed endonucleasi AP viene aggiunto alle cellule irradiate in coltura, il danno UV può essere eliminato.

AGOSTO 2009 Chiudi finestra Ivan il Terribile, Pietro il Grande e Yaroslav il Saggio. Il suo soprannome è il Granduca di Kiev...

Perché le persone parlano lingue diverse? Non esiste una risposta definitiva a questa domanda. Gli studiosi e la Bibbia la interpretano diversamente....

Italia fino alla fine dell'XI sec. sopravvisse a diverse conquiste, a seguito delle quali fu divisa in tre regioni con differenti ...
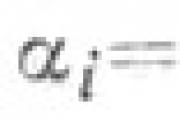
Definizione Una figura geometrica costituita da tutti i punti di un piano racchiusi tra due raggi emanati da uno ...

Entrare qui è il sogno di tanti giovani uomini e donne. Concorso annuale tra i candidati - più di 100 persone per posto. Di loro...

Se hai mai surfato o nuotato, probabilmente conosci una delle qualità più potenti dell'oceano: il suo...

È importante scegliere la giusta angolazione, regolare la luce se possibile, mascherare le imperfezioni, enfatizzare...

È noto che l'acqua è lo standard per convertire la massa in volume e viceversa, il volume in massa. Anche uno studente...

Qual è la bevanda analcolica più famosa al mondo? Certo, Coca Cola! Tutti lo percepiamo come qualcosa...

La limonata era fatta con succo di agrumi, acqua e zucchero. Tutti hanno bevuto questa bevanda analcolica con piacere ...

Beeline ha annunciato oggi che disabiliterà BlackBerry Internet Service per smartphone basati su BBOS dal 21...

OGE 2017. Tedesco. Un insieme di materiali per la preparazione degli studenti. Vetrinskaya V.V. M.: 20 17. - 9 6 p. +...

Dall'autore Progettazione tematica 170 ore (5 ore settimanali) Progettazione tematica 136 ore (4 ore settimanali)...
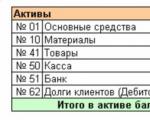
Il lavoro di contabilità di ogni contabile si basa sull'uso della logica e della matematica, richiede un ampio ...

Kelly Rohrbach è una modella e attrice americana, protagonista del film Baywatch. Infanzia e giovinezza Kelly Rohrbach ...

Per umiliare moralmente una persona, non basta trovare poche frasi su Internet, memorizzarle e ...