Vyhliadky na produkciu systémov transfúzie krvi
Newchemistry: Aké transfúzne systémy vyrábate? JSC "Syntéza", Kurgan: Vyrábame systémy pre ...
Minerálna výživa rastlín
Organizmus je v súlade s biotopom, z ktorého prijíma energiu a substráty, ktoré potrebuje pre svoju životne dôležitú činnosť, aby vybudoval a dynamicky aktualizoval biologické štruktúry. Táto príručka pojednáva o forme chemických prvkov v prostredí rastliny ao spôsoboch a spôsoboch ich vstupu do tela rastliny. Rastliny minerálnej výživy - je proces asimilácie minerálnych solí iónov z vonkajšieho prostredia, ktoré sú nevyhnutné pre normálne fungovanie rastlinného organizmu.
Prvky minerálnej výživy rastlín zahŕňajú predovšetkým makronutrienty N, P, S, K, Ca, Mg, ako aj stopové prvky (Fe, B, Cu, Zn, Mn, atď.). Tento zoznam prvkov neobsahuje kyslík, uhlík ani vodík. Je to spôsobené skutočnosťou, že uhlík je dodávaný v dostatočnom množstve z atmosférického zásobníka vo forme oxidu uhličitého - CO2, zatiaľ čo vodík vstupuje do tela rastliny v dôsledku rozkladu vody, zatiaľ čo uvoľnený kyslík sa uvoľňuje do životného prostredia. Poruchy minerálnej výživy. pozostáva z absorpcie minerálnych látok vo forme iónov, ich pohyb cez rastlinu a začlenenie do metabolizmu . Jednobunkové organizmy a vodné rastliny absorbujú ióny po celom povrchu a vyššie suchozemské rastliny absorbujú povrchové koreňové bunky, najmä vlasové korene uvedené vyššie.
Ióny sa najprv adsorbujú na bunkové membrány, potom prenikajú do cytoplazmy cez okolitú lipoproteínovú membránu - plazmatickú membránu. Katióny (s výnimkou K +) prenikajú membránou pasívne, difúziou, aniónmi a K + (pri nízkych koncentráciách) - aktívne, s použitím molekulárnych „iónových čerpadiel“, ktoré transportujú ióny s energetickým výdajom. Rýchlosť transportu aktívnych iónov závisí od dostupnosti sacharidov v bunke a intenzity dýchania, rýchlosť pasívnej absorpcie závisí od priepustnosti biologických membrán, rozdielu koncentrácií a elektrických potenciálov medzi médiom a bunkou. Priepustnosť membrány pre rôzne ióny sa mení. Takže pre K + katión je 100-krát vyšší ako pre Na + a 500 krát vyšší ako pre anióny.
Absorbované ióny sa pohybujú z bunky do bunky prostredníctvom cytoplazmatických mostov, ktoré ich spájajú - plazmidov. Vyššie rastliny v koreňoch a stonkách majú špeciálny cievny systém na prepravu minerálnych látok a ich organických zlúčenín (ktorých syntéza sa čiastočne vyskytuje v koreňoch) do listov. Ako dolné listy starnú, niektoré minerály prúdia z nich do rastúcich rastlinných orgánov, kde môžu byť znovu použité.
Každý prvok minerálnej výživy rastlín hrá určitú úlohu v metabolizme a nemôže byť úplne nahradený iným prvkom. Dusík je súčasťou proteínov - hlavných látok cytoplazmy, ako aj zloženia amidov, nukleových kyselín, hormónov, alkaloidov, vitamínov (B 1, B 2, B 6, PP) a chlorofylu. Dusík sa absorbuje vo forme aniónu NO - 3 (nitrát) a katiónu NH + 4 (amónia), ktoré vznikajú pri rozklade podstielky pôdnymi mikroorganizmami.
Molekulový dusík (N2), ktorý je hlavnou zložkou vzduchu (79%), môže byť asimilovaný iba určitými druhmi nižších rastlín. . Dusičnany sa redukujú na amónium pomocou enzýmu nitrát reduktázy. Amoniak sa kombinuje s organickými kyselinami za vzniku aminokyselín, ktoré sú potom včlenené do proteínov.
Fosfor je súčasťou nukleoproteínov bunkového jadra, fosfolipidov bunkové membrányfosfatidy a fosfátové estery cukrov. Mimoriadne dôležitá je účasť fosforu na fotofosforylácii, pri ktorej sa slnečná energia akumulovaná vo forme energeticky bohatých adenozín trifosfátových väzieb (ATP) používa na absorbovanie CO2 zo vzduchu a tvorbu organických látok. Vo forme makroergických väzieb ATP tiež ukladá energiu uvoľnenú počas dýchania v dôsledku oxidácie organických látok. , vznikli v procese fotosyntézy .
Fosfor sa absorbuje vo forme aniónu kyseliny ortofosforečnej (PO 3-4 alebo fosfát) a je zahrnutý pre stotiny sekundy v organických zlúčeninách v nezmenenej forme. Rastliny však vždy obsahujú veľa anorganického fosfátu (jeho fyziologický význam nie je jasný).
Síra, podobne ako dusík, je súčasťou všetkých proteínov, ako aj peptidov (glutatión), niektorých aminokyselín (cystín, cysteín, metionín) a éterických olejov. Síra je absorbovaná rastlinami vo forme aniónu (S02-4 alebo sulfátu), ktorý je redukovaný v bunkách, pričom vytvára disulfidové (-S-S-) a sulfhydrylové (-SH) skupiny (tieto tvoria väzby, ktoré fixujú konfiguráciu proteínovej makromolekuly). Síra spravidla nie je nedostatočným prvkom pre rastliny, ktoré nepotrebujú ďalšie kŕmenie zlúčeninami síry.
Draslík sa absorbuje vo forme katiónu K + a zostáva v bunke v rovnakej forme, bez tvorby silných organických zlúčenín. Vstupuje len do slabých adsorpčných interakcií s proteínmi a výmennými reakciami s organickými kyselinami. Na rozdiel od N, P a S, ktoré sa priamo podieľajú na tvorbe organického materiálu rastlinnej bunky, draslík nie je plná živina. Zvyšuje schopnosť cytoplazmy zadržiavať vodu, intenzitu fotosyntézy, tok asimilátov, podieľa sa na fungovaní žalúdka a pod.
Vápnik a horčík sa absorbujú vo forme dvojmocných katiónov: Ca2 + a Mg2 +. Hlavnou funkciou Ca je stabilizácia bunkových štruktúr. Ióny Ca2 + („vápnikové mostíky“) viažu lipidové molekuly spoločne a zaisťujú ich usporiadané usporiadanie v bunkových membránach. Zlúčeniny vápnika s pektínovými látkami zlepujú membrány susedných buniek. Na rozdiel od iných prvkov rastlinnej minerálnej výživy má Ca v rastline nízku mobilitu. Prakticky sa nepoužíva a akumuluje v starnúcich orgánoch. Ca je nevyhnutný na udržanie štruktúry ribozómu. , v ktorých dochádza k syntéze proteínov. Mg je súčasťou chlorofylu, aktivuje enzýmy, ktoré prenášajú fosfát z ATP na molekulu cukru. Železo je súčasťou radu enzýmov, vrátane respiračného (cytochrómu). . Podieľa sa na syntéze chlorofylu, hoci nie je zahrnutý v jeho zložení. Je možné aj minerálnu výživu rastlín cez listy.
Spolu s výživou ovzdušia (fotosyntéza) predstavuje minerálna výživa rastlín jediný proces metabolizmu medzi rastlinou a prostredím. Ovplyvňuje všetky fyziologické procesy (dýchanie, rast, vývoj, fotosyntéza, vodný režim, atď.) A závisí od nich. Jedným z najúspešnejších spôsobov riadenia produktivity obrábaných rastlín je preto regulácia minerálnej výživy rastlín a jej optimálne udržiavanie. To sa zvyčajne vykonáva s hnojivom.
Úloha dusíka vo výžive rastlín
V živom organizme je dusík jednou z hlavných živín, ktoré tvoria najdôležitejšie látky živých buniek - proteínov a nukleových kyselín. Množstvo dusíka v tele je však relatívne malé - od 1 do 3% sušiny. Relatívny obsah dusíka v materskej skale - hlavná oblasť tohto prvku - atmosféra - je zanedbateľný. Iba niektoré mikroorganizmy a modrozelené riasy môžu absorbovať molekulárny dusík v atmosfére, výrazné zásoby dusíka sa koncentrujú v pôde vo forme rôznych minerálov (amónne soli, dusičnany) a organických zlúčenín (dusík proteínov, nukleových kyselín a produktov ich rozkladu, teda nie celkom rozložených zvyškov rastlín a živočíchov).
Rastliny absorbujú dusík z pôdy vo forme anorganických a niektorých organických zlúčenín. Za prirodzených podmienok sú pre výživu rastlín veľmi dôležité pôdne mikroorganizmy (amoniakátory), ktoré mineralizujú dusík organickej pôdy na amónne soli. Dusičnanový dusík v pôde vzniká v dôsledku vitálnej aktivity otvoreného SN Vinogradského v roku 1890 nitrifikačných baktérií oxidujúcich amoniak a amónne soli na dusičnany. Časť dusičnanového dusíka absorbovaného mikroorganizmami a rastlinami sa stráca a mení sa na molekulárny dusík pod vplyvom denitrifikačných baktérií.
Rastliny a mikroorganizmy môžu absorbovať amónny i dusičnanový dusík, čím sa redukujú na amoniak a amónne soli. Mikroorganizmy a rastliny aktívne konvertujú anorganický amónny dusík na organické zlúčeniny dusíka - amidy (asparagín a glutamín) a aminokyseliny.
Ako ukázali D. N. Pryanishnikov a V.S. Butkevich, dusík v rastlinách sa skladuje a transportuje vo forme asparagínu a glutamínu. Tvorba týchto amidov neutralizuje amoniak, ktorého vysoké koncentrácie sú toxické nielen pre zvieratá, ale aj pre rastliny. Amidy sú súčasťou mnohých proteínov, a to ako v mikroorganizmoch, tak aj v rastlinách, ako aj vo zvieratách. Glutamín a asparagín sa syntetizujú enzymatickou amidáciou kyseliny glutámovej a asparágovej nielen v mikroorganizmoch a rastlinách, ale aj v rámci určitých limitov u zvierat.
Syntéza aminokyselín sa uskutočňuje redukčnou amináciou množstva aldehydových kyselín a ketokyselín vznikajúcich oxidáciou sacharidov (V. L. Kretovich) alebo enzymatickou transamináciou (A.E. Braunshteyn a M. G. Kritzman, 1937). Konečnými produktmi asimilácie amoniaku mikroorganizmami a rastlinami sú proteíny, ktoré tvoria protoplazmu a bunkové jadro, ako aj ukladané vo forme skladovacích proteínov.
Zvieratá a ľudia môžu syntetizovať iba v obmedzenom rozsahu aminokyseliny. Nemôžu syntetizovať 8 esenciálnych aminokyselín (valín, izoleucín, leucín, fenylalanín, tryptofán, metionín, treonín, lyzín), a preto pre nich hlavným zdrojom dusíka sú proteíny spotrebované z potravy, t. J. rastlín a mikroorganizmov.
Proteíny vo všetkých organizmoch podliehajú enzymatickému rozkladu, ktorého konečnými produktmi sú aminokyseliny. V nasledujúcom stupni, ako výsledok deaminácie, sa organický dusík aminokyselín opäť prevedie na anorganický amónny dusík. V mikroorganizmoch a najmä v rastlinách sa môže amóniový dusík použiť na novú syntézu amidov a aminokyselín. U zvierat sa neutralizácia amoniaku, ktorý vzniká počas rozkladu proteínov a nukleových kyselín, uskutočňuje syntézou kyseliny močovej (u plazov a vtákov) alebo močoviny (u cicavcov, vrátane ľudí), ktoré sa potom z tela odstránia.
Z hľadiska metabolizmu dusíka rastliny sú jednak zvieratá a ľudia na druhej strane rozdielne v tom, že zvieratá využívajú len slabý amoniak - väčšina z nich je eliminovaná z tela; v rastlinách je výmena dusíka „uzavretá“ - dusík, ktorý vstupuje do zariadenia, sa vracia do pôdy len so samotnou elektrárňou.
Konverzie dusíka v pôde
Organické zlúčeniny obsahujúce dusík tvorené v rastlinách prostredníctvom trofických reťazcov vstupujú do tela heterotrofov, ako aj do pôdy ako podstielky, po smrti rastlín a zvierat. V pôde prechádzajú rozkladom za účasti saprofágov, sú mineralizované a potom používané inými rastlinami. Rozklad konečného spoja sú organizmy - amoniakátory, tvoriace amoniak (NH3). Amoniak je zahrnutý do nitrifikačných reakcií, t.j. tvorby dusitanov a ich premeny na dusičnany. Cyklus cyklu dusíka v pôde sa teda udržiava neustále.
Súčasne sa časť dusíka vracia do atmosféry v dôsledku aktivity baktérií - denitrifikátorov, ktoré rozkladajú dusičnany na molekulárny dusík (N2). V dôsledku denitrifikácie baktérií sa z 1 ha pôdy ročne vyparí až 50-60 kg dusíka.
Zloženie sušiny rastlín, obsah dusíka nie je príliš veľký, od 1 do 3%. Avšak dusík je súčasťou proteínov a nukleových kyselín - hlavných biopolymérov bunky, bez ktorých život nie je možný. V pôde je viazaný dusík reprezentovaný štyrmi typmi zlúčenín: dusík amónne soli (NH 4 +); dusičnanový dusík (NO 3 -); organický dusík proteínov a nukleových kyselín vo forme degradačných zvyškov rastlín a živočíchov, ako aj ich produkty štiepenia - aminokyseliny, peptidy, amíny a amidy, ako aj dusík humusu. Je známe, že anorganické formy dusíka-amónneho a dusičnanového dusíka sú oveľa lepšie absorbované rastlinami ako jeho organické zlúčeniny. Výnimkou sú látky, z ktorých sa ľahko delí amónny dusík - močovina, asparagín a glutamín. Je to pôdne mikroorganizmy, ktoré mineralizujú organický dusík v pôde a premieňajú ho na amoniak - počiatočnú zlúčeninu, ktorú rastliny pripájajú na sacharidy a tvoria aminokyseliny a proteíny.
Cyklus dusíka
Všeobecným smerom biogeochemického cyklu dusíka na planéte je jeho akumulácia v molekulárnej forme N2 v atmosfére, približne 78%. Živá hmota a pôda sú však proti tomuto trendu. Biosféra obsahuje približne 150 miliárd ton dusíka, viazaných v organických zlúčeninách pôdneho pokryvu - 1,5 · 10 11 ton, v rastlinnej biomase - 1,1.10 9 ton a živočíšnej biomase - 6,1 · 10 7 ton.
Dusík je súčasťou mnohých organických zlúčenín, najmä bielkovín. V molekule proteínu tvorí silné amidové väzby s uhlíkom alebo sa kombinuje s vodíkom, pričom je prítomný ako amínová alebo amidová skupina. Tvorba amidových (peptidových) väzieb (C-N-väzieb) je hlavným mechanizmom pre syntézu proteínových molekúl a peptidov, ktoré tvoria podstatu všetkého života na Zemi.
Schéma odrážajúca cyklus dusíka je znázornená na obr.

Obrázok 15 - Schéma cyklu dusíka. Hlavné kroky sú zvýraznené a sú uvedené odhady množstva dusíka v hlavných prúdoch. Čísla v zátvorkách - teragramy (Tr = 10 6 ton) za rok (podľa Y. Odumu, 1986)
Zdrojom dusíka pre autotrofy sú dusičnany (soli kyseliny dusičnej HNO 3), ako aj atmosférický molekulárny dusík. Dusičnanový dusík cez koreňový systém rastlín vstupuje do ciest v listoch, kde sa používa na syntézu rastlinných proteínov.
Druhým spôsobom, akým dusík vstupuje do tela, je priama fixácia dusíka z atmosféry. Tento fenomén je úplne unikátny a charakteristický pre prokaryoty - mikroorganizmy bez jadra. Do roku 1950 boli známe iba tri taxóny mikroorganizmov, ktoré sú schopné viazať atmosférický dusík:
Voľne žijúce baktérie rodov Azotobacter a Clostridium;
Symbiotické uzlové baktérie rodu Rhizobium
Modrozelené riasy (cyanobaktérie) rodov Anabaena, Nostoc, ako aj iní členovia radu Nostocales.
Potom boli objavené ďalšie typy organizmov, ktoré sú schopné fixovať dusík z atmosféry; purpurové baktérie rodu Rhodospirilum, ako aj pôdne baktérie v blízkosti Pseudomonas, aktinomycet z nodov koreňov olše (Alnus, Ceanothus, Myrika a ďalšie). Bolo tiež zistené, že modrozelené riasy rodu Anabaena (treba zdôrazniť, že tieto riasy majú schopnosť heterotrofného kŕmenia a majú iné príznaky, ktoré možno pripísať baktériám s rovnakým úspechom ako baktérie) môžu byť symbiózy húb, machov, papradí a dokonca semien a schopnosť fixovať dusík je prospešná pre oboch účastníkov. Táto schopnosť je dôvod, že pri pestovaní ryže a strukovín na rovnakom poli niekoľko rokov môžete získať dobré výnosy, bez toho, aby sa dusíkaté hnojivá.
Biochemický mechanizmus priamej fixácie atmosférického dusíka sa uskutočňuje za účasti enzýmu dusíka, ktorý katalyzuje rozklad molekuly dusíka (N2). Tento proces vyžaduje značné množstvo energie na prerušenie trojitej väzby v molekule dusíka. Reakcia prebieha za účasti molekuly vody, v dôsledku čoho vzniká amoniak (NH3), napríklad v uzlinách strukovín, na fixáciu 1 g dusíka, baktérie spotrebujú približne 10 g glukózy (približne 40 kcal) syntetizovanej počas fotosyntézy, t.j. je len 10%, tento proces bude podrobnejšie diskutovaný neskôr.
Uvedený príklad tiež ilustruje prínosy symbiózy ako stratégie „spolupráce“, ktorá podporuje prežitie. Nie je ťažké dospieť k myšlienke vyhliadky na pestovanie takýchto odrôd poľnohospodárskych plodín, ktoré by s využitím symbiózy s mikroorganizmami, ktoré fixujú dusík, poskytovali dobré výnosy bez použitia hnojív.
K suspenzii dusíkového cyklu môže dôjsť v dôsledku jeho akumulácie v hlbokých oceánskych sedimentoch. Súčasne sa dusík odstaví z okruhu niekoľko miliónov rokov. Straty sú kompenzované prúdom plynného dusíka počas sopečných erupcií. Y. Odum verí, že sopečné erupcie sú v tomto zmysle užitočné, a ak „blokujú všetky sopky na Zemi, potom viac ľudí môže zomrieť hladom, než teraz trpia erupciami“.
Cyklus dusíka je príkladom dobre pufrovaného cyklu plynných látok. Je to dôležitý faktor obmedzujúci alebo kontrolujúci počet organizmov. Cyklus dusíka bol podrobne študovaný. Je známe najmä to, že z 10 9 ton dusíka, ktoré sa každoročne asimilovali v biosfére, sa približne 80% vracia do obehu z pôdy az vody a iba 20% požadovaného množstva je „nový“ dusík z atmosféry s dažďom a v dôsledku fixácie dusíka. Naopak, z dusíka vstupujúceho do polí s hnojivami sa opätovne používa veľmi malá časť; veľká časť sa stráca pri zbere v dôsledku odstraňovania vody a denitrifikácie.
V prirodzených biogeocenózach sa vďaka vyrovnávajúcim zložkám fytocenózy vytvára nulová bilancia dusíka. Poskytuje sa procesmi, ako je rozklad rastlinných zvyškov a humusu, syntéza organických látok de novo, amoniakácia, denitrifikácia, fixácia dusíka.
V agrocenózach, keď je biologická rovnováha v ekosystéme prudko narušená, je situácia úplne iná: po rozoraní panenských pôd klesá fixácia dusíka a vyvíja sa negatívna bilancia dusíka. Poľnohospodárstvo na Sibíri, kde sa monokultúra pšenice bez hnojiva často pestuje v starých agrocenózach, vykazuje pozitívnu bilanciu dusíka. Skutočnosť, že plodina v týchto rokoch dosahuje v priaznivých rokoch 20 centov na hektár, dokazuje, že sa tu uskutočňuje aktívna fixácia dusíka, čo čiastočne kompenzuje stratu dusíka v dôsledku zberu a denitrifikácie.
Úloha fosforu vo výžive rastlín
Fosfor je základným prvkom výživy rastlín. Je súčasťou nukleových kyselín, membrán, fosfolipidov. Fosfor je súčasťou energetického systému, je súčasťou vysokoenergetických zlúčenín. Ako je skladovacia látka uložená v semenách rastlín. Ak minerálna výživa nemá fosfor, potom aktivita fotosyntézy a respirácie klesá, pretože je narušená syntéza chlorofylu.
Už dlho sa pozorovalo, že v prvých obdobiach rastu plodiny absorbujú fosfáty intenzívnejšie ako v nasledujúcich. Hladina fosforu v rastlinách v počiatočnom období rastu spôsobuje taký dlhý depresívny účinok, že ho nemožno úplne prekonať ani normálnou následnou výživou. Okrem toho takéto vyhladované kultúry na začiatku vývoja kultúry negatívne reagujú na bohatú výživu fosfátov v budúcnosti.
Problém fosforu vzniká jedným z najakútnejších v poľnohospodárstve. To je vysvetlené dvoma hlavnými dôvodmi - nedostatkom geologických zásob tohto prvku a jeho rýchlym a pevným viazaním v pôde pri aplikácii hnojivami. Z tohto dôvodu nie je stráviteľnosť fosforečných hnojív poľnohospodárskymi rastlinami vyššia ako 25% a jej obrovské množstvo je pevne stanovené v pôde, čo sa stáva pre fosfáty ťažké pre rastliny. .
Hrubé zásoby fosforu v pôde sú pomerne významné. Pestované pôdy môžu obsahovať 10–20 a dokonca 30 ton tohto prvku na hektár ornej vrstvy. Je však vo vode nerozpustnej forme, ktorá nie je pre rastliny ľahko dostupná. Hlavným zdrojom fosforu pre rastliny v prírodných podmienkach sú soli kyseliny ortofosforečnej.
Kyselina trifosforečná ortofosforečná môže disociovať za vzniku troch aniónov: H2P04 -, HPO4 - a PO4 - -. V podmienkach slabo kyslej reakcie pôdy, v ktorej sú rastliny najčastejšie pestované, je najčastejšie prvá, ale druhá z uvedených iónov je tiež zastúpená.
Všetky soli jednoväzbových katiónov (NH4 +, Na +, K +) a kyseliny ortofosforečnej vyskytujúce sa v pôde sú dobre rozpustné vo vode a ľahko sa absorbujú poľnohospodárskymi plodinami. Soli monovalentných katiónov a kyseliny metafosforečnej sú tiež rozpustné. Fosfáty dvojmocných katiónov (Ca2 +, Mg2 +) sú rozpustné vo vode len v prvom stupni substitúcie v kyseline ortofosforečnej a slabo rozpustné aj v tejto substitúcii v kyseline metafosforečnej.
Disubstituované soli dvojmocných katiónov a kyseliny ortofosforečnej sú nerozpustné vo vode, ale rozpustné v slabých kyselinách, vrátane organických, uvoľňovaných koreňmi do pôdy a mikroorganizmov v nej obsiahnutých počas životnej aktivity. Z týchto dôvodov sú disubstituované soli katiónov dvojmocnej kyseliny ortofosforečnej dôležitým zdrojom stráviteľného fosforu pre rastliny.
Trisubstituované fosfáty dvojmocných katiónov vo väčšine poľnohospodárskych rastlín sú oveľa ťažšie stráviteľné, pretože tieto soli sa vyznačujú nerozpustnosťou vo vode a veľmi obmedzenou rozpustnosťou v slabých kyselinách a potom väčšinou v čerstvo vyzrážanej forme, keď sú v amorfnom stave. So stárnutím solí sa rozpustnosť prudko znižuje a dostupnosť fosforu pre rastliny sa znižuje.
Existuje však skupina kultúr schopných živiť sa fosforom a v prítomnosti len troch substituovaných fosforečnanov vápenatých alebo prírodných fosforitov v jemne mletej forme. Patrí medzi ne vlčí bôb, pohánka, horčica, espartit, ďatelina, hrach a konope. Schopnosť týchto rastlín kŕmiť fosforečnany nerozpustné v fosfore je primárne spôsobená kyslosťou vylučovania koreňov. Bolo zistené, že v roztoku okolo koreňových chĺpkov lupiny je pH 4 - 5 av podobnej bazálnej zóne ďateliny je to 7 - 8.
Fosforový cyklus
Zásobník fosforu, na rozdiel od dusíka, nie je atmosférou, ale horninami a inými usadeninami, ktoré vznikli v minulých geologických epochách. Minerálny fosfor je súčasťou mnohých hornín. Vstupuje do hydrosféry počas ich hypergenézy, ukladá sa vo forme sedimentov v plytkých vodách a čiastočne uložený v hlbokomorských sedimentoch.
U zvierat je súčasťou kostí a iných tkanív fosfor vo forme organických zlúčenín (najmä proteínov). Tiež hrá úlohu v energetických procesoch uskladnenia bunkovej energie vo forme adenozíntrifosfátu a adenozín difosforečných kyselín. V dôsledku rozkladu odumretých organizmov a mineralizácie organických zlúčenín sa fosfor vo forme fosforečnanov (soli kyseliny ortofosforečnej) opäť používa v rastlinách a tým sa opäť zúčastňuje cirkulácie.
Odstránenie fosforu z obehu nastáva v dôsledku jeho akumulácie v spodných sedimentoch. Cyklus fosforu je príkladom jednoduchého sedimentárneho cyklu s nedostatočným „vztlakom“ a narušenými mechanizmami samoregulácie v dôsledku antropogénneho vplyvu na životné prostredie. Existuje názor, že mechanizmy návratu fosforu do obehu sú nedostatočné a nevyvažujú straty spojené s technogenézou.
Ľudská činnosť v oblasti rybolovu a vtákov vedie k nerovnováhe fosforu. Podľa J. Hutchinsona, len asi 60.000 ton elementárneho fosforu sa vracia do pôdy v dôsledku rybolovu. Každý rok sa na hnojivá vyťaží 1 až 2 milióny ton hornín obsahujúcich fosfor a väčšina tohto množstva sa vymyje vodou a odstráni sa z okruhu. V súčasnosti vzbudzuje obavy zvýšenie koncentrácie fosfátov vo vodných ekosystémoch, čo vedie k ich intenzívnemu prerastaniu, degradácii ekosystémov a nakoniec k ich smrti.
Fosfor sa široko používa v poľnohospodárskej technológii vo forme fosfátových (minerálnych) hnojív s cieľom zvýšiť úrodnosť pôdy a výnosy plodín. Minerálny fosfor tak vstupuje do vodných a suchozemských ekosystémov v dôsledku odstraňovania rozpustených fosfátov z poľnohospodárskej odpadovej vody a odtoku z oblastí, v ktorých sa používajú fosfátové hnojivá, ako aj vypúšťania mestských a priemyselných odpadových vôd.
Podľa J. Hutchinsona je doba obratu fosforu vo vode malých jazier (0,3 - 0,4 km 2 a hĺbka 6 - 7 m) 5,4 - 7,6 dní a veľké (2 km 2, hlboké 2 km 2). asi 4 m) - 17 dní. Doba obratu v dnových sedimentoch je oveľa dlhšia a predstavuje približne 40 a 176 dní. Rozdiel v hodnote ukazovateľa je zrejme spôsobený tým, že v malých jazerách je pomer povrchu spodných sedimentov k objemu vody väčší. Vo veľkých, ale nie hlboko-vodných nádržiach sa tak ukladá fosfor, čo značne komplikuje boj proti ich prerastaniu.
Úloha draslíka vo výžive rastlín
Draslík v tele je jednou z najdôležitejších živín, trvalej zložky rastlín a živočíchov. Mnohé morské organizmy získavajú draslík z vody. Rastliny si ju z pôdy. Na rozdiel od sodíka sa draslík koncentruje hlavne v bunkách, jeho extracelulárne médium je mnohokrát menšie. Draslík je tiež nerovnomerne distribuovaný v bunke.
Ióny draslíka podporujú osmotický tlak a hydratáciu koloidov v bunkách, aktivujú niektoré enzýmy. Metabolizmus draslíka. Súvisia s metabolizmom sacharidov, ióny K + ovplyvňujú syntézu proteínov. K + vo väčšine prípadov nemožno nahradiť Na +. Bunky selektívne koncentrujte K +. Inhibícia glykolýzy, respirácie, fotosyntézy, zhoršenej permeability vonkajšej bunkovej membrány vedie k uvoľneniu K + z buniek, často výmenou za Na +.
Enzýmy fosforfruktokinázy a pyruvátkinázy, ktoré sa podieľajú na prenose fosforečnanových zvyškov bohatých na energiu, tiež vyžadujú na preukázanie svojej aktivity draslíkový katión. Draslík zvyšuje aktivitu amylázy, sacharázy a proteolytických enzýmov. Nedostatok vedie k dezorganizácii metabolizmu v rastlinnom tele.
Zdá sa, že v rastline je draslík v iónovej forme. V žiadnom prípade nie sú známe organické zlúčeniny, ktoré sú syntetizované v organizmoch, ktorých súčasťou by bol draselný katión. Najmenej 80% je v bunkovej miazgeli rastlín a extrahuje sa vodou. Menšia časť draslíka je adsorbovaná koloidmi a približne 1% je absorbované mitochondriami v protoplazme. Je obsiahnutý hlavne v protoplazme a vakuolách. V jadre a plastidoch nie je draslík.
V rastlinách je draslík tiež nerovnomerne rozdelený: vo vegetatívnych orgánoch rastliny je viac ako v koreňoch a semenách. Veľa draslíka v strukovinách, repách, zemiakoch, tabakových listoch a trávnych trávach (20-30.) g / kgsušiny). S nedostatkom draslíka v pôde spomaľuje rast rastlín, zvyšuje výskyt. Draslík zlepšuje kvalitu poľnohospodárskych výrobkov: zvyšuje sa akumulácia cukrov v cukrovej repe a škrob v zemiakových hľúzach. V druhom prípade je kyselina sírová draslíka účinnejšia. U ľanu a konope sa zvyšuje výťažok a kvalita vlákniny v obilninách, zvyšuje sa celková hmotnosť zŕn a zvyšuje sa hmotnosť 1000 zŕn. Hodnota draslíka a jeho hlavné funkcie v živote rastlín sú znázornené na obr.
S hladom draslíka sa znižuje odolnosť zemiakov, zeleniny a cukrovej repy voči hubovým chorobám, a to tak počas obdobia rastu, ako aj počas čerstvého skladovania. S nedostatkom draslíka v obilninách sa slama stáva menej trvanlivou, chlieb je položený, čo vedie k zníženiu úrody, zhoršuje plnenie obilia. vstup hnojivo zvyšuje obsah vo vode rozpustných foriem draslíka v pôde, zabraňuje vzniku koreňovej hniloby (Helminthosporiumsativum) a znižuje infekčný potenciál pôdy.
Rôzne plodiny konzumujú nerovné množstvo draslíka. Konzumujte relatívne veľa ovocie a bobuľoviny, cukrovej repy, kapusty, koreňovej zeleniny, zemiakov, ďateliny, lucerny, slnečnice, pohánky, kukurice a strukovín. Na vytvorenie zrna je potrebných menej draslíka. Na rozdiel od dusíka a fosforu je draslík väčší vo vegetatívnom ako v reprodukčných orgánoch rastlín (semená).

Obrázok 16 - Hlavné funkcie draslíka v rastlinnom živote
Napríklad, v slame pšenice ozimnej, raže, draslíka jačmeňa takmer
Pri správnom a úplnom použití organického odpadu sa draslík vracia do pôdy vo väčších množstvách ako dusík a fosfor. Na vytvorenie optimálnej potašovej výživy pre rastliny s vysokým obsahom dusíka a fosfátov je však spravidla potrebné aplikovať do pôdy priemyselné hnojivá. Draselná pôda je jej hlavným zdrojom výživy rastlín. Jeho celkový obsah v pôde často presahuje obsah dusíka a fosforu. To je do značnej miery určené povahou rodičovského plemena.
Tabuľka 5 - Priemerný obsah draslíka vo výťažku najdôležitejších plodín,% z celkovej hmotnosti
|
kultúra |
obilia |
slama |
kultúra |
obilia |
slama |
|
Zimná pšenica |
0,50 |
0,90 |
sója |
1,26 |
0,50 |
|
Zimná raž |
0,60 |
1,00 |
vika |
0,80 |
0,63 |
|
kukurica |
0,37 |
1,64 |
Široké fazuľa |
1,29 |
1,94 |
|
jačmeň |
0,55 |
1,00 |
Modrý vlčí bôb |
1,14 |
1,77 |
|
ovos |
0,50 |
1,60 |
Ľan (semená) |
1,00 |
0,97 |
|
proso |
0,50 |
1,59 |
Alfalfa (seno) |
1,50 |
|
|
pohánka |
0,27 |
2,42 |
Clover (seno) |
1,50 |
|
|
hrášok |
1,25 |
0,50 |
V biosfére sú stopové prvky Rb a Cs sprevádzané draslíkom a sú schopné ich nahradiť, pričom ich koncentrácia je nedostatočná. Ionty Li + a Na + sú antagonisty K +, preto sú dôležité nielen absolútne koncentrácie K + a Na +, ale aj optimálne pomery K + / Na + v bunkách a médiu. Prírodná rádioaktivita organizmov (žiarenie gama) o takmer 90% je spôsobená prítomnosťou prírodného rádioizotopu 40 K v tkanivách.
Draslík, spolu s dusíkom a fosforom, je jedným z najdôležitejších prvkov minerálnej výživy v poľnohospodárstve. Rýchlosť potaše závisí od typu poľnohospodárskeho hnojiva. plodín a pôdy.
Stav draslíka v pôde
Jeho kôra obsahuje 2,14%. Nie menej sa to deje v sedimentárnych horninách, ktoré sú materské pre mnohé pôdy. Množstvo draslíka v pôde je určené hlavne distribúciou veľkosti častíc. V ílovitých a hlinitých pôdach dosahuje jeho obsah 2% alebo viac - príloha B. Je to spôsobené tým, že v ťažkých pôdach je súčasťou minerálov, zastúpených najmä v ílovitých časticiach. Výrazne menej draslíka v piesočnatých, piesčitých a najmä rašelinových pôdach. Jeho množstvo v týchto pôdach je znížené na 0,1%.
Vzhľadom na to, že sa veľkosť častíc zväčšuje, zvyšuje sa obsah draslíka. To možno ilustrovať príkladom podzolizovanej ťažkej hlinitej pôdy agrochemickej experimentálnej stanice Dolgoprudnaya. DN Pryanishnikov a obyčajný hlinitý chernozem Ústavu poľnohospodárstva Strednej čiernej pôdy. VV Dokuchaeva - tabuľka 6.
Tabuľka 6 - Obsah draslíka v jednotlivých frakciách distribúcie veľkosti častíc pôdy,%
Draslík ílovitej frakcie je pre rastliny najprístupnejší, pretože je obsiahnutý hlavne vo výmennom stave. Hrubý obsah draslíka v pôde zriedkavo charakterizuje dostupnosť rastlín pre nich, pretože v pôde je len asi 1% hrubých rezerv dostupných pre rastliny. Preto by sa dostupnosť rastlín s draslíkom na rôznych pôdach nemala posudzovať podľa celkového percenta jeho obsahu v pôde, ale podľa pomeru medzi formami jej zlúčenín. Hrubý obsah draslíka v podložnej vrstve pôdnej podzolovej pôdy a prechodný je často vyšší ako v ornej pôde - Tabuľka 7.
Podľa dostupnosti rastlín možno všetky zlúčeniny draslíka v pôde rozdeliť do piatich skupín:
Draslík rôznych pôdnych minerálov, hlinitokremičitanov. Táto forma obsahuje najvyššie množstvo draslíka. Je to skôr v orthokláze, menej v muskovite, biotite, glaukonite, nefelíne a leukute. Táto forma draslíka je pre rastliny ťažká. V roku 1947 sa z pôdy izolovali baktérie nazývané silikátové baktérie schopné rozkladať ortoklázu. Niektorí výskumníci sa domnievajú, že zohrávajú pozitívnu úlohu v výžive rastlín draslíka. Predpokladá sa, že časť ortoklázy draslíka a leucitu ako minerálov, ktoré sú najviac odolné voči kyselinám, môžu byť premenené na prístupnú formu v dôsledku mykorhizie niektorých trvalých plodín.
Draslík muskovit, biotit a nefelín sú dostupnejšie pre rastliny. Časť z nich ide do asimilácie rastlín ako výsledok výmenného rozkladu so soľami pôdneho roztoku. Určité množstvo draslíka týchto minerálov sa môže stať prístupným v dôsledku pôsobenia oxidu uhličitého a niektorých organických kyselín emitovaných koreňmi rastlín na nich.
Tabuľka 7 - Obsah draslíka v ornej a podpovrchovej vrstve rôznych pôd,%
|
pôda |
Ukážkové umiestnenie |
||
|
A (na ornej pôde) |
B (na ornej pôde) |
||
|
Horská tundra |
Hibiny |
2,87 |
3,37 |
|
SOD-podzoly |
Leningradský región |
3,10 |
3,78 |
|
Šedá lesná step |
Tula |
2,81 |
3,07 |
|
černozeme |
Kamenná step (Voroněžská oblasť) |
2,64 |
2,13 |
|
Ľahký gaštan |
Severný Kaukaz |
2,67 |
2,38 |
|
serozem |
Územie Stavropolu |
2,34 |
2,14 |
|
červená pôda |
okres Batumi (Adjara) |
0,52 |
0,22 |
V závislosti od typu pôdy prebieha prechod draslíka z nezmeniteľných do vymeniteľných foriem s rôznou intenzitou. Na pôdach pod-podzolic sú tieto hodnoty 15–30 kg / ha ročne, na vylúhovanom černozeme - približne 60 kg / ha.
Draselné koloidy pôdy. Táto forma - hlavný zdroj výživy draslíka rastlín. V pôde to môže byť 5 - 30 mg / 100 g. Jeho množstvo v pôde ako percentuálny podiel z celkového obsahu závisí od typu a podtypu pôdy, najmä od distribúcie veľkosti častíc. Napríklad na piesočnatých pôdach je táto forma draslíka iba 0,8%, na hlinitej pôde - 1,5 a na čierno-zemej a šedej pôde - 1 - 3%.
Podiel draslíka na množstve absorbovaných báz nemôže indikovať stupeň bezpečnosti rastlín. Napríklad podľa K.K. V ornej vrstve (0 - 20 cm) hlinitého cereínu (oblasť Tula) bol draslík 2,7%, v podzolizovanej hlinke (Smolensk) 6,1% z celkového množstva absorbovaných báz. Absorpčná kapacita v prvom prípade bola 54,8, v druhej - 6,21 mmol100 g pôdy. Preto výmena draslíka v čiernej pôde bola 70,65, v podzolizovanej hlinke iba 17,9 mg / 100 g pôdy.
Jedným z najdôležitejších úloh agrochémie je stanoviť stupeň účasti zameniteľného draslíka pôdy pri tvorbe plodiny. Rastliny používajú počas vegetačného obdobia len časť vymeniteľného draslíka, v závislosti od vlastností pôdy, biologických vlastností rastlín a poveternostných podmienok. Objektívne údaje o potrebách rastlín na hnojivá z potaše možno získať chemickými metódami a stanovením experimentov s hmotnostným poľom v špecifických pôdnych a klimatických podmienkach. Malá časť draslíka (1 - 5 mg / kg pôdy) je v pôdnom roztoku vo forme solí uhlia, kyseliny dusičnej, fosforečnej, sírovej, chlorovodíkovej a iných kyselín.
Vo vode rozpustný draslík. Obsah tejto formy prvku je 1 / 5–1 / 10 časti množstva K2O, ktorý je v pôde vo výmennom stave. V ornej vrstve černozemu je to asi 0,02–0,06 mmol / 100 g pôdy, v alkalických pôdach - 0,08–0,10, v sodno-podzolovom - 0,04–0,09 mmol / 100 g pôdy. Roztok pôdy zriedka obsahuje viac ako 0,1 mmol (4,7 mg) K20 / 100 g pôdy. V neúspešnej pôde podzolovej pôdy TSCA, počas jarno-letného obdobia, sa množstvo draslíka rozpustného vo vode pohybovalo od 1,5 do 5 mg / kg pôdy alebo 4,5-18 kg / ha.
Vo vode rozpustný draslík je najviac dostupný pre výživu rastlín. Vyskytuje sa v pôde najmä vďaka chemickým a biologickým účinkom na pôdne minerály, ako aj ich hydrolýze. Napríklad minerály môžu byť zničené pod vplyvom koreňových výlučkov rastlín, kyslé odpadové produkty mikroorganizmov, vrátane kyseliny dusičnej, nahromadené nitrifikačnými baktériami. Časť draslíka môže prejsť z výmenného stavu do roztoku v dôsledku jeho vytesnenia z absorpčného komplexu rôznymi soľami, vrátane hnojív zavedených do pôdy.
Draslík, ktorý je súčasťou plazmatických mikroorganizmov. V pôde s podzolovou pôdou dosahuje množstvo 40 kg K 2 O
1 ha. Tento draslík vstupuje do dostupnej formy až po smrti mikróbov. Nesmieme však zabúdať, že spolu s procesom odumierania sa mikroorganizmy rýchlo množia. A to si vyžaduje všetky živiny, vrátane draslíka. Preto je ťažké posúdiť, koľko draslíka sa uvoľňuje, keď mikroorganizmy umierajú a sú dostupné pre rastliny, a tiež absorbované v procese ich reprodukcie. Tieto otázky sú stále zle pochopené. Draslík sa nachádza aj v rastlinných, živočíšnych, koreňových a rastlinných zvyškoch, hnojoch a iných organických látkach vstupujúcich do pôdy. Po ich rozklade sa stáva dostupným pre rastliny.Draslík fixovaný v pôde. V pôde nielen transformácia draslíka z ťažko rozpustných foriem na vymeniteľné a vo vode rozpustné formy, ale aj spôsoby fixácie draslíka v nezmenenom stave, t. upevnenie pôdy. Tento proces prebieha aktívne s premenlivým zmáčaním a sušením pôdy. Pôda s veľkou distribúciou veľkosti častíc obsahujúca veľký počet jemných frakcií sa vyznačuje zvýšenou fixáciou draslíka. Draslík je zvlášť účinný v prítomnosti ílových minerálov montmorillonitovej skupiny a hydromiky v pôde, ktoré sú charakterizované intrakryštalickou adsorpciou katiónov. Kaolínová skupina ílových minerálov nemá túto vlastnosť.
Rôzne typy pôd majú nerovnomernú schopnosť fixovať draslík v nezmenenom stave. Najintenzívnejšie draslík je fixovaný v slonkách. Predpokladá sa, že peptizácia spôsobená alkalizáciou zvyšuje počet koloidných častíc v ílových mineráloch a tým podporuje vstup draslíkových katiónov do ich kryštálovej mriežky. Chernozems opraviť draslík lepšie ako sod-podzolic pôdy.
Zvýšené množstvo organickej hmoty v pôde, ako aj vápnenie kyslých pôd, posilňuje fixáciu draslíka v nezmenenej forme. Systematické nanášanie potašových hnojív znižuje fixáciu draslíka pôdou, pretože fixačná kapacita pôdy nie je nekonečná. Zo všetkých katiónov, ktoré sú dôležité pre výživu rastlín, sú amónne a draselné pevné. Upevnenie jedného z týchto prvkov zabraňuje a dokonca zabraňuje upevneniu druhého.
Uzamykacia schopnosť pôdy sa prejavuje do určitého limitu. Fixácia draslíka pôdou drasticky znižuje jeho využitie z aplikovaných hnojív. Napríklad na pochodových (aluviálnych) pôdach Holandska sa zaznamenáva 21 - 59% draslíka aplikovaného počas mnohých rokov. V Kanade sa v dôsledku fixácie draslíka pôdou rastliny použilo len 25–48% tohto prvku, aplikovaného minerálnymi hnojivami.
Väčšina fixného draslíka je v ornici. Systematické používanie hnojív zvyšuje obsah rôznych foriem draslíka v porovnaní s nepohodlnými možnosťami. Povaha transformácie draslíka však vo veľkej miere závisí od pôdnych a klimatických podmienok. V pôde s podzolovými a sivými lesmi sa značne zvyšuje množstvo vymeniteľného draslíka. Napríklad v pôdach agrochemickej experimentálnej stanice Dolgoprudnaya počas 36 rokov sa zvýšila o 8-10 mg / 100 g pôdy. Obsah nezmeniteľného draslíka sa mierne zvýšil, čo je vysvetlené absenciou podmienok pre jeho fixáciu (nadmerná vlhkosť, nízka teplota, kyslá reakcia atď.).
Výmena draslíka pri systematickej aplikácii hnojív na tieto pôdy sa akumuluje nielen na ornej pôde, ale aj v hlbších vrstvách. Vzhľadom na vysokú saturáciu dvojmocnými katiónmi, černozem takmer neakumuluje vymeniteľný draslík. Nevýhodná absorpcia draslíka v dôsledku priaznivých podmienok na jeho fixáciu (zloženie ílových minerálov, absencia umývacieho režimu, veľké množstvo organických látok atď.) Prevláda. Zvýšenie množstva nezmeniteľného draslíka sa zvyčajne pozoruje v ornej a podložnej vrstve pôdy a dosahuje významné hodnoty.
Systematické hnojenie sierozem vedie k výraznému zvýšeniu obsahu vymeniteľných a nezmeniteľných draslíka. Zavlažovanie prispieva k akumulácii výmenného a nezmeniteľného draslíka pozdĺž profilu pôdy do hĺbky 1 m.
Fixácia draslíka z hnojív na sodno-podzolických pôdach je malá a zriedka presahuje 200 kg / ha. Absorpcia draslíka dosahuje v chernozme značnú veľkosť a dosahuje 300 - 700 kg K 2 O na 1 ha. Čím kratšia je dĺžka používania hnojív a množstvo zavedeného draslíka, tým väčšia je jeho relatívna fixácia. Je zrejmé, že použitím vysokých dávok potašových hnojív na chernozem je možné dosiahnuť úplné nasýtenie ich fixačnej kapacity a bez strachu z fixácie draslíka použiť v súčasnosti akceptovanú metódu periodického aplikovania potašových hnojív.
Úloha stopových prvkov vo výžive rastlín
K stopovým prvkom patria chemické prvky prítomné v organizmoch v nízkych koncentráciách (zvyčajne jedna tisícina percenta alebo menej). Termín stopové prvky sa používa na označenie určitých chemických prvkov obsiahnutých v pôdach, skalách, mineráloch a vode. Presné kvantitatívne kritériá na rozlíšenie mikroprvkov od makroprvkov neboli stanovené. Niektoré makroživiny pôdy a hornín (Al, Fe, atď.) Sú stopové prvky pre väčšinu zvierat, rastlín a ľudí.
V živých organizmoch sa jednotlivé stopové prvky našli už na začiatku 19. storočia, ale ich fyziologický význam zostal neznámy. V. Vernadsky zistil, že mikroprvky nie sú náhodnými zložkami živých organizmov a že ich distribúcia v biosfére je určená množstvom zákonitostí. Podľa moderných údajov viac ako 30 mikroelementov. považované za nevyhnutné pre život rastlín a zvierat. Väčšina stopových prvkov sú kovy (Fe, Cu, Mn, Zn, Mo, Co a ďalšie), niektoré sú nekovy (I, Se, Br, F, As).
V tele sa stopové prvky nachádzajú v rôznych biologicky aktívnych zlúčeninách: enzýmy (napríklad Zn - v karboanhydráze, Cu - v polyfenol oxidáze, Mn - v argináze, Mo - v xantínoxidáze, približne 200 metaloenzýmov), vitamíny (Co - vitamín B 12), hormóny (I - na tyroxín, Zn a Co - na inzulín), respiračné pigmenty (Fe - na hemoglobín a iné pigmenty obsahujúce železo, Cu - na hemocyanín). Pôsobenie stopových prvkov, ktoré tvoria tieto zlúčeniny alebo ovplyvňujú ich funkciu, sa prejavuje najmä zmenami aktivity metabolických procesov v organizmoch.
Niektoré stopové prvky ovplyvňujú rast (Mn, Zn, I - u zvierat; B, Mn, Zn, Cu - v rastlinách), reprodukciu (Mn, Zn - u zvierat; Mn, Cu, Mo - v rastlinách), tvorbu krvi (Fe, Cu, Co), na procesoch tkanivového respirácie (Cu, Zn), intracelulárneho metabolizmu atď. Pre množstvo stopových prvkov nájdených v organizmoch (Sc, Zr, Nb, Au, La a niektoré ďalšie) nie je ich kvantitatívna distribúcia v tkanivách a orgánoch známa. a biologická úloha nie je jasná.
Stopové prvky v pôdach sú súčasťou rôznych zlúčenín, ktorých významnú časť tvoria nerozpustné alebo ťažko rozpustné formy a len malá časť mobilnými formami, ktoré môžu rastliny absorbovať. Mobilita stopových prvkov a ich dostupnosť pre rastliny je vo veľkej miere ovplyvnená kyslosťou pôdy, vlhkosťou, obsahom organických látok a ďalšími podmienkami, ako aj aktivitou mikroorganizmov.
Obsah stopových prvkov v pôdach rôznych typov sa líši. Napríklad chernozémy sú bohaté na mobilné formy B a Cu (0,4-1,5 a 4-30 mg v 1 kilogrampôda) a chudobná sod-podzolic (0,02-0,6 a 0,1-6,7 mg v 1 kilogram), mo je deficientný v pľúcach, Co v kyslej sodno-podzolovej pôde, Mn v chernozmoch, Zn v hnedých a gaštanových pôdach. Nedostatok alebo nadbytok stopových prvkov v pôde vedie k ich nedostatku alebo nadbytku v rastlinnom a živočíšnom tele. Súčasne dochádza k zmenám v povahe akumulácie, oslabeniu alebo posilneniu syntézy biologicky aktívnych látok, reorganizácii procesov intersticiálneho metabolizmu, vzniku nových adaptácií alebo vzniku porúch vedúcich k tzv. Endemickým ochoreniam ľudí a zvierat.
Napríklad endemická ataxia u zvierat je spôsobená nedostatkom Cu, nadbytkom Mo a sulfátov, prípadne aj Pb; endemická struma u ľudí a zvierat - nedostatok I; acobalt - nedostatkom Co v pôde; Borická enteritída, komplikovaná pneumóniou (u oviec), je prebytok B. V rôznych biogeochemických provinciách endemické choroby zvyčajne postihujú 5-20% hospodárskych zvierat v obci - x. zvierat alebo populácií druhov. Nedostatok alebo nadbytok stopových prvkov je tiež škodlivý pre rastliny.
Takže s nedostatkom Mo sa potlačuje tvorba kvetov v karfiole av niektorých strukovinách; s nedostatkom Cu - je narušená tvorba ovocia v obilninách, citrusových a iných rastlinách; s nedostatkom B - nedostatočne rozvinutej nádoby, nie je kvitnúce (arašidy), púčiky (jablko, hruška) zomrieť, kvetenstvo (hrozno) a ovocie (arašidy, kapusta) vyschnúť; s nadbytkom V rastlinách sú postihnuté hnilobou koreňového goliera, ochorejú pri chloróze, vznik škvŕn sa rozširuje. V provinciách, kde koncentrácia jednotlivých mikroprvkov nedosahuje dolné prahové hodnoty, je možné predísť endemickým chorobám a ich vyliečenie pridaním príslušných mikroprvkov do krmiva pre zvieratá, mikroživiny sa používajú pre rastliny. .
Stopové prvky sú v tele nerovnomerne rozložené. Ich zvýšená akumulácia v jednom alebo inom orgáne je do značnej miery spojená s fyziologickou úlohou prvku a špecifickou aktivitou orgánu (napríklad prevládajúca akumulácia Zn v pohlavných žľazách a jeho vplyv na reprodukčnú funkciu); v ostatných prípadoch mikroelementy ovplyvňujú orgány a funkcie, ktoré nesúvisia s miestom ich akumulácie v tele.
Hnojivá v poľnohospodárskej výrobe
Hnojivá zahŕňajú organické a anorganické látky, ktoré obsahujú rastlinné živiny alebo mobilizujú živiny v pôde. V závislosti od chemického zloženia sú rozdelené na organické hnojivá (hnoj, komposty, zelené hnojivá atď.) A minerálne hnojivá (dusíkaté, fosforečné, potašové, komplexné, vápenaté, mikronutričné hnojivá) a priame a nepriame hnojivá. Existuje aj skupina bakteriálnych hnojív. .
Hnojivá vyrobené priamo na farmách sú klasifikované ako lokálne a tie, ktoré sa vyrábajú v chemických závodoch, sú priemyselné. Hnojivá zvyšujú úrodnosť pôdy (jej režimy výživy, vody, tepla a vzduchu), zlepšujú chemické, fyzikálno-chemické, fyzikálne a mikrobiologické vlastnosti. Opakovaná aplikácia hnojív vo veľkých dávkach a iných metódach obrábania pôdy (spracovanie, výsadba bylín, atď.) Môže zmeniť smer procesu tvorby pôdy a viesť k vzniku nových pôdnych podtypov - antropogénnych pôd s vysokou plodnosťou.
Aplikovaním hnojív človek aktívne zasahuje do obehu látok v prírode, pričom vytvára najmä pozitívnu bilanciu živín v poľnohospodárstve. Keď sa hnojivá používajú správne, majú pozitívny vplyv na rast, vývoj rastlín a napokon aj na výnos a kvalitu produktu. Účinnosť hnojív závisí od biologických vlastností obce. plodín, obsah živín v pôde a jej vlhkosť, reakcia pôdneho roztoku atď. Veľké zvýšenie výnosu dáva hnojivo pod zavlažovanie. Kombinácia hnojív a zavlažovanie tiež prispieva k hospodárnejšiemu využívaniu vody a živín rastlinami. Dostupnosť hnojív na hektár plodín je jedným z hlavných ukazovateľov intenzifikácie poľnohospodárstva.
Produkcia a spotreba, ako aj tempo rastu hnojív neustále rastú, ak porovnáte tieto hodnoty z tabuľky 50 - 70 s tabuľkou 8 s dynamikou týchto ukazovateľov, posledné desaťročia, znázornené na obrázku 17, ukazujú stály rast ich výroby a spotreby.
Tabuľka 8 - Svetová produkcia minerálne hnojivá (Tisíc. t účinnej látky - N, P205 a K2O)
|
hnojivá |
||||
|
leta |
dusík (N) |
fosforečná (P 2 O 5) |
potaš (K 2 O) |
celkom (NPK) |
|
1950 |
4382 |
6120 |
4315 |
14817 |
|
1955 |
7106 |
8719 |
6915 |
22740 |
|
1960 |
10564 |
10703 |
8668 |
29935 |
|
1965 |
18788 |
15669 |
12678 |
47135 |
|
1970 |
31911 |
21286 |
17564 |
70761 |
|
1972 |
36060 |
23906 |
19795 |
79761 |

Obrázok 17 - Miera spotreby a rastu hnojív
Obrázok 18 znázorňuje štruktúru spotreby hnojív v roku 2006 podľa odhadov IFA. Ako vyplýva z grafu, viac ako polovica spotrebovaných minerálnych hnojív dusíkatých hnojív - 59%. Podiel fosfátových hnojív predstavuje štvrtinu vyrobených hnojív - 24%, výroba hnojív na výrobu potaše predstavuje 17%.

Obrázok 18 - Štruktúra spotreby hnojív v roku 2006 (odhad IFA) (na základe materiálov z internetu)
azotnye hnojivo
Dusíkaté hnojivá sú minerálne a organické látky používané ako zdroj dusíkovej výživy rastlín. Rozdeľujú sa na organické hnojivá (hnoj, rašelina, kompost), ktoré okrem dusíka obsahujú aj iné rastlinné živiny a minerálne hnojivá vyrábané v priemysle, ako aj zelené hnojivo alebo zelené hnojivá (vlčí bôb, serodella atď.).
Dusíkaté hnojivá sa používali už v staroveku. Pri obrábaní rôznych krajín sa široko používal hnoj. V zavlažovanom poľnohospodárstve Strednej Ázie je už dlho známe zelené hnojivo. Oveľa neskôr začali používať minerálne hnojivá, z ktorých prvý bol dusičnan sodný, ťažený od polovice 19. storočia. z prírodných ložísk v Čile (Južná Amerika). Jeho spotreba v roku 1900 bola asi 300 tisíc. t(z hľadiska dusíka). V nasledujúcich rokoch priemysel začal vyrábať síran amónny, kyánamid vápenatý a dusičnan vápenatý.
Do roku 1913 dosiahla celosvetová produkcia dusíkatých hnojív takmer 700 tisíc. t – 700*10 3 t (z hľadiska dusíka). Priemyselný vývoj syntézy amoniaku z dusíka zo vzduchu a vodíka (1914–18) umožnil prudký nárast svetovej produkcie, ktorá sa v roku 1966 zvýšila na 19 200 tis. t(z hľadiska dusíka).
Od roku 2000 sa celosvetová spotreba dusíkatých hnojív od roku 2000 zvýšila o 12% a v roku 2005 predstavovala 90,86 mil. Ton - 90,96 * 10 6 t, Najvýraznejší nárast spotreby dusíkatých hnojív v rokoch 2000 - 2005 zabezpečovali krajiny južnej a východnej Ázie - 18,4%, resp. 22,2%. V posledných troch rokoch spotreba v západnej Európe a Severnej Amerike poklesla.
V roku 2006 sa zvýšenie celosvetovej spotreby dusíkatých hnojív odhaduje na 2,7%, čo je o niečo menej v porovnaní s inými druhmi minerálnych hnojív. Hlavnou hnacou silou dopytu po hnojivách podľa prognóz IFA zostane Ázia. Južná a východná Ázia poskytne odhadovanú polovicu celkového rastu spotreby. V budúcnosti, do roku 2010, IFA očakáva zvýšenie priemerného ročného nárastu spotreby dusíkatých hnojív - o viac ako 1,8% v dôsledku ich nahradenia komplexnými hnojivami a ich efektívnejšieho využívania. V nasledujúcich rokoch IFA odhaduje nadbytok ponuky hnojív nad dopytom, ktorý by v roku 2010 mohol dosiahnuť 22,5 mil. Ton - obrázok 19.

Obrázok 19 - Miera spotreby a rastu dusíkatých hnojív (podľa internetu)
V minerálnych dusíkatých hnojivách môže byť dusík vo forme amoniaku (NH3), amoniaku-dusičnanu (NH3 a N03), nitrátu (NO3) a amidu (NH2). Amoniak hnojivá zaujímajú dôležité miesto medzi dusíkatými hnojivami. Amónne hnojivá zahŕňajú síran amónny, chlorid amónny, hydrogenuhličitan amónny, kvapalné hnojivá na báze amoniaku. Síran amónny a chlorid amónny sú najúčinnejšie na pôdach nasýtených zásadami (chernozem, uhličitan sierozem, gaštan), ktoré majú schopnosť neutralizovať okysľujúci účinok týchto hnojív.
Systematické hnojenie síranom amónnym a chloridom amónnym v kyslých pôdach spôsobuje zvýšenie kyslosti; tento nedostatok môže byť eliminovaný vápnením. Dusík amoniak je menej náchylný na vylúhovanie ako dusičnanový dusík, takže hnojivá na báze amónia sa môžu aplikovať pred výsevom na jeseň. Sú menej vhodné na zavedenie povrchu (pri kŕmení zimných plodín) a miestneho (v radoch, dierach a hniezdach). Nadbytok chlóru v chloride amónnom negatívne ovplyvňuje veľkosť a kvalitu mnohých poľnohospodárskych plodín. plodiny (zemiaky, ľan, olejniny, tabak, hrozno atď.). Hydrogenuhličitan amónny má alkalickú reakciu, ale v pôde sa podrobí nitrifikácii. Medzi amoniakovými formami hnojív na báze amoniaku sú veľmi dôležité kvapalné hnojivá - kvapalný bezvodý amoniak, vodný amoniak a amoniaky.
Hnojivá na báze dusičnanu amónneho: dusičnan amónny a(dusičnan amónny, dusičnan amónny), sulfonitrát amónny (dusičnan pruhu, dusičnan montánny, nitrosulfát amónny). Dusičnan amónny sa vyrába hlavne v granulovanej forme; slabo okyslí pôdu. Sulfonitrát amónny má relatívne vysokú schopnosť okysľovania.
Nitrátové hnojivá - dusičnan sodný (dusičnan sodný, dusičnan sodný, dusičnan čilský), dusičnan vápenatý (dusičnan vápenatý, dusičnan vápenatý, dusičnan vápenatý, nórsky dusičnan), dusičnan draselný (dusičnan draselný, dusičnan draselný). Dusičnan sodný je fyziologicky alkalické hnojivo, preto je lepšie ho aplikovať na kyslé pôdy, najmä v cukrovej repe, pšenici, jačmeni a iných plodinách citlivých na kyslosť pôdy. Dusičnan vápenatý sa vyrába v granulovanej forme, zvyčajne s prímesou dusičnanu amónneho; tiež alkalizuje pôdu. Dusičnan draselný okrem dusíka obsahuje draslík a je zdrojom dusíkovo-draselnej výživy rastlín. Prineste ho pod kultúru citlivú na chlór. Všetky dusičnanové formy dusíka nie sú absorbované v pôde. V oblastiach s nadmernou vlhkosťou na ľahkých pôdach so slabou kapacitou zadržiavania vody sa vylúhujú dusičnanové hnojivá, preto sa ako hlavné hnojivo odporúča používať hnojivá na báze čpavku.
Amidové hnojivá - močovina (karbamid), kyánamid vápenatý, močovinoformaldehydové hnojivá. Najcennejšia močovina. V pôde sa ľahko zmení na uhličitan amónny; najprv trochu alkalizuje a potom mierne okyslí pôdu. Kyánamid vápenatý má schopnosť znižovať kyslosť pôdy. Účinné na sypkých, organicky bohatých neutrálnych pôdach, ak sú na jeseň oplodnené. Nevhodné pre lokálnu aplikáciu. Kyánamid vápenatý sa tiež používa ako defoliant na odstraňovanie listov z bavlny pred zberom. Močovinoformaldehydové hnojivá sa z pôdy nevypierajú; sú obzvlášť účinné v oblastiach s nadmernou vlhkosťou a zavlažovaným poľnohospodárstvom. Môžete použiť vysoké dávky týchto hnojív, ktoré rastliny s dusíkom na niekoľko rokov. Charakteristiky minerálnych dusíkatých hnojív sú uvedené v tabuľke 9.
Tabuľka 9 - Vlastnosti základných minerálnych dusíkatých hnojív
|
hnojivá |
chemický vzorec |
priemerný dusík ( % ) |
objem závažia hnojivá (kg / m. *) |
hygroskopický nosta |
|
síran amónneho |
(NH4) 2S04 |
20,5-21,5 |
Veľmi slabý |
|
|
Chlorid amónny. |
NH4C1 |
26,0 |
chudobný |
|
|
Bezvodý amoniak. |
NH3 |
82,3 |
||
|
Amoniaková voda ...... |
NH3 + H20 |
20,0 |
||
|
Dusičnan amónny |
||||
|
granulovaný |
NH4NO3 |
34,7-35,0 |
Veľmi silný |
|
|
kryštál |
NH4NO3 |
34,7-35,0 |
Veľmi silný |
|
|
Dusičnan sodný |
NaNO3 |
16,0 |
1100 i400 |
umiernený |
|
Dusičnan vápenatý |
Ca (N03) 2 * 2H20 |
17,0 |
900-1100 |
Veľmi silný |
|
močovina |
||||
|
granulovaný |
(NH2) 2CO |
46,0 |
Veľmi slabý |
|
|
kryštál |
(NH2) 2CO |
46,0 |
Veľmi slabý |
Dusíkaté hnojivá sú účinným prostriedkom na zvyšovanie výnosov plodín, najmä v nepernozemi, vo vlhkých oblastiach lesostepu av zóne zavlažovaného poľnohospodárstva, kde pôdy obsahujú nedostatočné množstvo dusíka. Normy minerálnych dusíkatých hnojív závisia od pôdnych podmienok, biologických vlastností plodín, stupňa zabezpečenia hnojom alebo inými organickými hnojivami.
Približné miery dusíkatých hnojív (v kg na 1 ha z hľadiska dusíka): pre ozimné obilniny vysiate na rušnom páre, 40 - 60, na čistom páre 30 - 40; pod jarným zrnom 40 - 60; kukurica pre siláž a zrno v nepernozemskej zóne a severná časť lesostepnej zóny 60–120, na bohatej čerešňovej pôde-45–60, v zavlažovaných oblastiach 120–150; pod cukrovou repou na lesostepných chernozmoch 45–60, na sivej lesnej pôde, podzolizovaných chernozmoch lesokopy a v nepernozemskej zóne 80–120, v zavlažovaných oblastiach 100-150; pod bavlnou 120 - 140; ľanové dolgun 40 - 60; pod konope 45 - 90; so zemiakmi 45 - 90; pod kapustou 90 - 120; pod paradajkami, uhorkami 60 - 90; v prípade ovocných plodín 60 - 100%. \\ t
Menšie sadzby sa používajú na pôdach bohatších na prírodný dusík, ako aj na súčasné použitie hnoja alebo iných látok obsahujúcich dusík. organické hnojivo, S dostatočnými hladinami dusíka v pôde, v oblastiach s vlhkosťou, sa môžu zvýšiť množstvá hnojív, čo spravidla zvyšuje výnosy a zlepšuje kvalitu produktu. napríklad dobrá výživa dusíka podporuje tvorbu gluténu v pšeničnom zrne, zvyšuje obsah bielkovín v kŕmnych plodinách.
Dusíkaté hnojivá sa používajú ako hlavné hnojivo aj ako vrchné obväzy. Pod zimnými plodinami zasiatej na čistý pár, sú zavedené len v skorých jarných obväzov (30 - 40 kilogram dusíka na 1 hektár) na zamrznutej pôde. Pod jarnými plodinami vo všetkých zónach dusíkatých hnojív CIS. v plnej miere je vhodné použiť pred výsevom a na zavlažovanie v niekoľkých etapách pred zavlažovaním.
Fhnojivá
Jediným zdrojom dopĺňania zásob fosforu v pôde sú minerálne a organické látky obsahujúce fosfor a používané na zlepšenie výživy fosforu rastlín. Fosfátové hnojivá sa vyrábajú najmä priemyselne z banských surovín - fosfátovej horniny a apatitu. Organické látky ako kostná múčka, hnoj, troska bohatá na fosfor, rajčiaková troska a iné sa používajú aj ako fosfátové hnojivá.
Fosfátové hnojivá - prvé z minerálnych hnojív, vyrábané priemyselne. Superfosfát bol prvýkrát vyvinutý vo Veľkej Británii v roku 1842 - predtým sa ako fosfátové hnojivo používala hlavne kostná múčka.
Svetová produkcia fosfátových hnojív do roku 1900 predstavovala približne 1 milión. t (v zmysle P 2 O 5) . V 20. storočí (najmä zo stredu) ich použitie výrazne vzrástlo - tabuľka 10. Ich relatívny rast v poľnohospodárstve je však nižší ako rast dusíkatých a draselných hnojív, čo sa vysvetľuje nedostatočnými zásobami fosfátových surovín.
10. - Svetová spotreba v poľnohospodárstve fosforečných minerálnych hnojív, tis. t P205
|
krajiny |
1950 |
1960 |
1970 |
1974 |
|
Všetky krajiny vrátane: United States ZSSR Francúzsko Čína Austrália Nemecko Poľsko Japonsko Brazília India španielsko Kanada Veľká Británia Taliansko |
5918 1869 – |
9600 2427 1088 – |
18802 4145 3184 1684 |
24255 4600 4496 2152 1390 1171 |
Od roku 2000 sa celosvetová spotreba fosforečných hnojív zvýšila od roku 2000 o 13% av roku 2005 predstavovala 36,78 milióna ton. Výrazný nárast spotreby v rokoch 2000 - 2005 zabezpečili krajiny východnej a južnej Ázie a Latinskej Ameriky - 29,3%, 25% a 19%. V rovnakom období sa spotreba v krajinách západnej Európy, Oceánie a krajín WANEA znížila o 17,2%, 7,5% a 7,6%.
V budúcnosti, do roku 2010, IFA očakáva zvýšenie priemerného ročného rastu spotreby fosfátových hnojív o 2,6%. V nasledujúcich rokoch IFA odhaduje, že prebytok ponuky nad dopytom po hnojivách je stabilný na úrovni 1-2 miliónov ton. Hlavnými výrobcami fosfátových hnojív sú krajiny Severnej Ameriky a Ázie. Hlavnou surovinou na výrobu tohto typu hnojiva je koncentrát apatitu. Spotreba fosfátového hnojiva a rýchlosť rastu sú uvedené v grafe - obrázok 20

Obrázok 20 - Spotreba fosfátových hnojív a rýchlosť rastu
Podľa rozpustnosti sú fosfátové hnojivá rozdelené do 3 skupín. Vo vodorozpustných hnojivách (jednoduchý, dvojitý a amoniakálny superfosfát) je fosfor obsiahnutý vo forme monobázického fosforečnanu vápenatého Ca (P204) 2 * H20. Vyrábajú sa prevažne granulované a používajú sa v hlavnom a základnom nátere (v radoch). V hnojivách rozpustných v citráte av citrónoch rozpustných hnojivách (zrazenina, tomát, troska, fosfát, troska, defluorovaný fosfát, fosforečnan horečnatý) je fosfor vo forme dvojsýtneho fosforečnanu vápenatého CaHPO 4 H20 alebo fosforečnanu vápenatého Ca4P205. Tieto hnojivá sa používajú na hlavnú aplikáciu na orbu alebo kultiváciu.
V ťažko rozpustných hnojivách obsahujúcich fosfor - fosforečnanovú horninu, kostná múčka je fosfor obsiahnutý vo forme fosforečnanu vápenatého Ca3 (PO4) 2. Aplikujú sa ako hlavné hnojivo vo vysokých dávkach na kyslých pôdach, v ktorých sú pre rastliny dostupné slabo rozpustné fosfáty. Všetky fosfátové hnojivá nie sú hygroskopické, nespárujú sa a dobre sa rozprestierajú vŕtačkou.
Vysoko koncentrované fosfátové hnojivá sú sľubné - polyfosforečnany amónne, metafosforečnany draselné obsahujúce od 50 do 80% P205. Z hľadiska účinnosti sú ekvivalentné av niektorých prípadoch presahujú štandardné formy hnojív obsahujúcich fosfor. V Spojených štátoch a niektorých krajinách západnej Európy sa používajú kvapalné hnojivá na báze polyfosforečných kyselín. Použitie týchto hnojív umožňuje úplnú mechanizáciu ich aplikácie, zníženie strát na minimum, rovnomerné zapustenie pôdy a zároveň zavedenie stopových prvkov a pesticídov. Charakteristiky hlavných minerálnych fosfátových hnojív sú uvedené v tabuľke 11.
Tabuľka 11 - Charakteristiky hlavných minerálnych hnojív
|
hnojivá |
Chemický vzorec |
|
|
Superfosfát jednoduchý a zrnitý |
Sa (H20P04) 2H20 + 2CaS04 |
14–19,5 |
|
Dvojnásobne granulovaný superfosfát |
Sa (H20P04) 2x H20 |
|
|
Fosforečná múka |
CaF (PO4) 3 + CaOH (PO4) 3 + CaC03 |
19–30 |
|
vzniknutá zrazenina |
SaNPO 4 * 2H20 |
27–35 |
|
Fosfátová troska |
4СаО * Р 2 O 5 * СaSiO 3 |
16–19 |
|
Tomasshlak |
4СаО * Р 2 О 5 + 4СаО * P 2 O 5 * CaSiO 3 |
Fosfátové hnojivá zvyšujú výťažok a zlepšujú jeho kvalitu, urýchľujú zrenie rastlín, zvyšujú ich odolnosť voči usadzovaniu a suchu. Osobitne dôležité je to pre Kazachstan, kde sa hlavné poľnohospodárske oblasti nachádzajú v zóne s nedostatočnou vlhkosťou. Vysoká účinnosť fosfátových hnojív bola zavedená vo všetkých pôdno-klimatických pásmach krajiny, ak sa uplatňujú na všetky poľnohospodárske produkty. kultúre. Ich pozitívny účinok je zrejmý najmä v súvislosti s poskytovaním rastlín dusíku a draslíka.
Úvod 60 kilogramP 2 O 5 (základné hnojivo) pre ozimnú pšenicu dáva ďalších 2–5 u od 1 hektár obilia. V oblastiach pestovania jarnej pšenice 60–80 kilogram P205 zvyšuje výťažok o 1,5 - 2,5 u od 1 hektár. Kvôli nízkej mobilite F. y. majú následný účinok po dobu niekoľkých rokov: v suchých oblastiach po dobu 6-8 rokov, v zóne dostatočnej vlhkosti po dobu 2-3 rokov.
Dávky fosfátových hnojív závisia od pôdnych podmienok, kultúry a dostupnosti živín pre rastliny. V ZSSR sa ako hlavné hnojivo (na orbu alebo pestovanie) používa 60–120. kg / ha P 2 O 5 a osivo - 10–40 kg / ha P205. Krmný fosfor je spravidla neúčinný, s výnimkou zavlažovanej pôdy.
Na zavlažovaných územiach republík strednej Ázie a Azerbajdžanu použite 100–120 kg / haP205 pod bavlnou zvyšuje úrodu surovej bavlny o 3-5 uod 1 hektár. V zónach cukrovej repy 60–120 kg / haP205 zvyšuje výťažok cukrovej repy o 25-50 u od 1 hektár a zvýšiť obsah cukru v koreňových plodinách o 0,1–0,3%. S fosforovým hnojivom v dávke 90%. \\ T kg / ha výnosy zemiakov na sodno-podzolových a čerešňových pôdach sa zvyšujú pa 25-30 u od 1 hektár; obsah škrobu v hľúzach sa zároveň zvyšuje o 0,6–1,2%. Fosfátové hnojivá sú tiež účinné pri aplikácii na iné poľnohospodárske produkty. plodiny - krmivo, zelenina, ovocie
Potašové hnojivá
Na potaš hnojivá zahŕňajú minerály s vysoký obsah dostupný draslík, používaný ako zdroj výživy draslíka pre rastliny. Sú to zvyčajne vo vode rozpustné soli kyseliny chlorovodíkovej, sírovej a uhličitej, často s prímesou ďalších zlúčenín obsahujúcich draslík vo forme, ktorá je prístupná rastlinám.
Hlavným zdrojom hnojív potaše sú ložiská prírodných draselných solí, ktorých prvé veľké ložisko bolo objavené v Stassfurte v 40. rokoch. 19. storočie V nasledujúcich rokoch priemysel začal vyrábať chlorid draselný, dusičnan draselný, síran draselný atď. ložiská draselných solí boli objavené v mnohých krajinách. Do roku 1913 bola celosvetová produkcia potašových hnojív (z hľadiska K 2 O, v miliónoch) t) 1,19, v rokoch 1967 - 14,7, v rokoch 1970 - 19.
Po dobu piatich rokov od roku 2000 vzrástla svetová spotreba hnojív na výrobu potaše o 20% av roku 2005 predstavovala 26,44 milióna ton. Najvýraznejší nárast spotreby potašových hnojív v rokoch 2000 - 2005 zabezpečovali krajiny východnej a južnej, severovýchodnej a juhovýchodnej Ázie - 60,6%, 44,4% a 44,3%. V rovnakom období sa spotreba v západnej Európe, Severnej Amerike a Afrike znížila o 14,6%, 2,9% a 9,3%. V roku 2006 sa nárast svetovej spotreby hnojív na výrobu potaše odhaduje na 4%, čo je v porovnaní s inými druhmi minerálnych hnojív viac. V budúcnosti, do roku 2010, IFA očakáva zvýšenie priemerného ročného rastu spotreby hnojív na báze potaše o 3%. V nasledujúcich rokoch IFA odhaduje, že prebytok ponuky nad dopytom po hnojivách je stabilný na úrovni 6 miliónov ton - obrázok 21.

Obrázok 21 - Spotreba priemyselných hnojív a rýchlosť rastu
Draselné hnojivá sú rozdelené na surové draselné soli získané mechanickým spracovaním (triedenie, drvenie a mletie) prírodných draselných solí a koncentrovaných hnojív - chloridu draselného, síranu draselného, 30% a 40% draselných solí (zmes jemne mletého prírodného kainitu alebo sylvinitu). s chloridom draselným), ako aj potaš, magnézium draselný, elektrolyt draselný, popol atď. Charakteristiky základných hnojív na báze potaše sú uvedené v tabuľke 12.
Obvykle sa draselné hnojivá používajú na pozadí fosfátových alebo dusíkatých a fosfátových hnojív. Najvyššie výnosy sa zvyšujú na chudobných mobilných draslíkových pôdach: rašeliniskách, brehových, piesčitých a ľahkých hlinito-podzolických. V draslíkových hnojivách potrebujú rastliny aj sivé lesné hliny, podzolizované a vylúhované černozem a červené pôdy vlhkých subtropov (s dlhodobým pestovaním čajových rastlín a citrusových plodín).
Tabuľka 12 - Vlastnosti hlavných minerálnych draselných hnojív
|
umelé hnojivo |
Chemický vzorec |
nasiakavosť |
spekanie |
|
|
Chlorid draselný |
52-60 |
nápadný |
silný |
|
|
30% a 40% draselných solí |
KCl + m KCl3 * NaCl |
30-40 |
nepatrný |
nápadný |
|
Síran draselný |
K2S04 |
45-52 |
Veľmi slabý |
žiadny |
|
Silvinit |
M NaCl |
Najmenej 14. \\ T |
nepatrný |
nápadný |
|
kainit |
KCl * MgS04 * 3H20 |
8-12 |
chudobný |
To isté |
|
karnalit |
KCl * MgCl2 * 6H20 |
12-13 |
nepatrný |
To isté |
|
Kalimagne zia |
K2SO4 * MgS04 |
24-26 |
Veľmi slabý |
žiadny |
|
Elektrolyt draselný |
KCI a nečistoty |
nápadný |
nápadný |
Draslík je citlivejší na tie rastliny, ktoré konzumujú veľké množstvo tohto prvku: zemiaky, zelenina, cukrová repa, kŕmne korene, tabak, tabak. Ľan a konope, aj keď si z pôdy vyberú trochu draslíka, ale zle sa prispôsobia, takže ich pestovanie je spojené so zavedením hnojív potašu.
Draslík má pozitívny vplyv na kvalitu výrobkov: koreňové plodiny zvyšujú obsah cukru, zemiaky - obsah škrobu, zvlákňovacie kultúry - výnos a kvalitu vlákniny a kŕmne rastliny - obsah bielkovín (najmä na pozadí dusíkatých hnojív amónneho dusíka). Okrem toho, potaš hnojivá. zvýšiť odolnosť rastlín voči niektorým hubovým chorobám av zimnom období chlieb, strukoviny a viacročné výsadby zvyšujú odolnosť proti mrazu a suchu.
Účinnosť hnojív potaš závisí od obsahu príbuzných prvkov - sodíka, chlóru a ďalších. V prípade zemiakov, tabaku, hrozna, vlčí bôb - plodín citlivých na chlór - sa kvalita plodín zvyśuje zvyćajne len pri pridávaní dusićnanu draselného alebo uhličitanu draselného. Cukrová repa a niektoré ďalšie rastliny reagujú pozitívne na surový a zmiešaný sodík draselný. Preto v hlavných zónach pestovania repy sylvinit (obsahujúci okrem draslíka sodíka) výrazne zvyšuje výťažok koreňov a zvyšuje ich obsah cukru výraznejšie ako čistý chlorid draselný. Pod hroznom, pohánkou, tabakom, fazuľou, zemiakmi je nežiaduce pridávať surové draselné soli. Ak sa tieto plodiny oplodnia chloridom draselným, potom sa aplikujú len na jeseň na orbu, takže počas jesene a skorého jari sa chlór nevstrebaný do pôdy môže v podstate vylúhovať z ornice. Pre tieto kultúry je ešte lepšie použiť magnézium draselné, síran draselný, dusičnan draselný a popol. Použitie hnojív potašu, výhodne súčasne s fosfátovými hnojivami, zvyšuje produktivitu lúk a zlepšuje kvalitu sena.
Dávky potašových hnojív závisia najmä od pôdnych podmienok, fyziológie, vlastností hnojenej kultúry a vlastností hnojív. Na pôdnom podzolovom pôde prispievajú (K 2 O, v kg / ha) od 30 do 60 (pre konope sa dávka zvýši na 120), na sivej lesnej pôde a chernozems, 30-60 a 30-45 (pod konope, do 90), na červených zeminách a sierozmoch, 30-60.
Hnojivá s obsahom potaše sa zvyčajne používajú ako hlavné hnojivo na jeseň alebo na jar na orbu alebo kultiváciu. Hnojenie rastlín s draslíkom (ak to nebolo dostatočne vysadené na orbu) sa stalo rozšírené v pestovaní cukrovej repy, zemiakov, kukurice a niektorých rastlinných plodín. Je lepšie používať draslík spolu s dusíkom a fosfátové hnojivá, privádzajúc ich do uličky výživou rastlín do hĺbky najmenej 10-12 vidieť (vo výške 20 - 30%) kg / ha K20).
Malé zapustenie do pôdy s hnojivami z potaše, keď kŕmenie rastlín neposkytuje pozitívne výsledky. Preto je kŕmenie draslíkovej kultúry kontinuálneho výsevu (obilniny, strukoviny, byliny) nepraktické. Každý centr K 2 O, zavedený vo forme potašových hnojív, poskytuje v priemere nasledujúce zvýšenie výnosu (v \\ t u): surová bavlna 1-2, cukrová repa 35-40, zemiaky 20-33, ľanové vlákno asi 1,5, zrno zimy 3-5, jar 2-3, seno vysiatej trávy 20-33, lúčne seno 8-18. Trvanie aftereffect hnojív potaš je 3-4 roky.
Minerálna výživa rastlín, asimiláciu iónov minerálnych solí z vonkajšieho prostredia, ktoré sú nevyhnutné pre normálne fungovanie rastlinného organizmu. K prvkom M. p. zahŕňajú N, P, S, K, Ca, Mg a stopové prvky (Fe, B, Cu, Zn, Mn, atď.). M. str. pozostáva z absorpcie minerálnych látok vo forme iónov, ich pohybu cez rastlinu a začlenenia do metabolizmus. Jednobunkové organizmy a vodné rastliny absorbujú ióny po celom povrchu, vyššie suchozemské rastliny absorbujú povrchové bunky koreň, v podstate koreňové chĺpky. Ióny sa najprv adsorbujú na bunkové membrány, potom prenikajú do cytoplazmy cez okolitú lipoproteínovú membránu - plazmatickú membránu. Katióny (s výnimkou K +) prenikajú membránou pasívne, difúziou, aniónmi a K + (pri nízkych koncentráciách) - aktívne, s použitím molekulárnych „iónových čerpadiel“, ktoré transportujú ióny s energetickým výdajom. Rýchlosť transport aktívnych iónov závisí od poskytovania buniek sacharidmi a intenzity dýchania, rýchlosti pasívnej absorpcie - z priepustnosti biologických membrán, rozdiely v koncentráciách a elektrických potenciáloch medzi médiom a bunkou. Priepustnosť membrány pre rôzne ióny sa mení. Takže pre K + katión je 100-krát vyšší ako pre Na + a 500 krát vyšší ako pre anióny. Absorbované ióny sa pohybujú z bunky do bunky prostredníctvom svojich prepojených cytoplazmatických mostov - plasmodesmata. Vyššie rastliny v koreňoch a stonkách majú špeciálny cievny systém na prepravu minerálnych látok a ich organických zlúčenín (ktorých syntéza sa čiastočne vyskytuje v koreňoch) do listov. Ako dolné listy starnú, niektoré minerály prúdia z nich do rastúcich rastlinných orgánov, kde môžu byť znovu použité.
Každý prvok M. pp hrá určitú úlohu v metabolizme a nemôže byť úplne nahradený iným prvkom. Dusík je súčasťou proteínov - hlavných látok cytoplazmy, ako aj zloženia amidov, nukleových kyselín, hormónov, alkaloidov, vitamínov (B 1, B 2, B 6, PP) a chlorofylu. Dusík sa absorbuje vo forme aniónu NO - 3 (dusičnanu) a katiónu NH + 4 (amónia), ktorý vzniká pri rozklade humusu pôdnymi mikroorganizmami. Molekulový dusík (N 2), ktorý je hlavnou zložkou vzduchu (79%), môže byť asimilovaný iba určitými nižšími rastlinnými druhmi (pozri \\ t Mikroorganizmy fixujúce dusík ). Dusičnany sa redukujú na amónium pomocou enzýmu nitrát reduktázy. Amoniak sa kombinuje s organickými kyselinami za vzniku aminokyselín, ktoré sa potom inkorporujú do proteínov. Fosfor je súčasťou nukleoproteínov bunkového jadra, fosfolipidov bunkových membrán, fosfatidov a fosforečných esterov cukrov. Mimoriadne dôležitá je účasť fosforu na fotofosforylácii, pri ktorej sa slnečná energia akumulovaná vo forme energeticky bohatých adenozín trifosfátových väzieb (ATP) používa na absorbovanie CO2 zo vzduchu a tvorbu organických látok. Vo forme makroergických väzieb ATP tiež ukladá energiu uvoľnenú počas dýchania v dôsledku oxidácie organických látok (pozri Oxidačná fosforylácia ), v procese fotosyntéza. Fosfor sa absorbuje vo forme aniónu kyseliny ortofosforečnej (PO 3-4 alebo fosfát) a je zahrnutý pre stotiny sekundy v organických zlúčeninách v nezmenenej forme. Rastliny však vždy obsahujú veľa anorganického fosfátu (jeho fyziologický význam nie je jasný). Síra, podobne ako dusík, je súčasťou všetkých proteínov, ako aj peptidov (glutatión), niektorých aminokyselín (cystín, cysteín, metionín) a éterických olejov. Síra je absorbovaná rastlinami vo forme aniónu (S02-4 alebo sulfátu), ktorý je redukovaný v bunkách, pričom vytvára disulfidové (-S-S-) a sulfhydrylové (-SH) skupiny (tieto tvoria väzby, ktoré fixujú konfiguráciu proteínovej makromolekuly). Draslík sa absorbuje vo forme katiónu K + a zostáva v bunke v rovnakej forme, bez tvorby silných organických zlúčenín. Vstupuje len do slabých adsorpčných interakcií s proteínmi a výmennými reakciami s organickými kyselinami. Na rozdiel od N, P a S, priamo zapojených do tvorby organického materiálu rastlinnej bunky, K nie je v plnom zmysle živinou. Zvyšuje schopnosť cytoplazmy zadržiavať vodu, intenzitu fotosyntézy, odtok asimilátov, podieľa sa na fungovaní žalúdka, atď. Vápnik a horčík sa absorbujú vo forme dvojmocných katiónov: Ca2 + a Mg2 +. Hlavnou funkciou Ca je stabilizácia bunkových štruktúr. Ióny Ca2 + („vápnikové mostíky“) viažu lipidové molekuly spoločne a zaisťujú ich usporiadané usporiadanie v bunkových membránach. Zlúčeniny Ca s pektínovými látkami spolu zlepujú škrupiny susedných buniek. Na rozdiel od iných prvkov M. pp, Ca v rastline má nízku mobilitu. Prakticky sa nepoužíva a akumuluje v starnúcich orgánoch. Ca je potrebné na udržanie štruktúry. ribozómy, v ktorých dochádza k syntéze proteínov. Mg je súčasťou chlorofylu, aktivuje enzýmy, ktoré prenášajú fosfát z ATP na molekulu cukru. Železo je súčasťou radu enzýmov, vrátane respiračných ( cytochrómami ). Podieľa sa na syntéze chlorofylu, hoci nie je zahrnutý v jeho zložení. Je možné aj M. p. cez listy (pozri Listová výživa rastlín ).
Spolu s dodávkou vzduchu ( fotosyntéza ) M. p. P. je jediný proces metabolizmu medzi rastlinou a prostredím. Ovplyvňuje všetky fyziologické procesy (dýchanie, rast, vývoj, fotosyntéza, vodný režim, atď.) A závisí od nich. Jedným z najúspešnejších spôsobov riadenia produktivity pestovaných rastlín je preto regulácia M. p. pomocou hnojív.
: Pryanishnikov D.N., Agrochemistry, Fav. cit., zv. 3, M., 1952; Kursanov AL, Vzájomný vzťah fyziologických procesov v rastline, M., 1960; Kolosov I. I., Absorpčná aktivita rastlinných koreňových systémov, M., 1962; Sutcliffe JF, Absorpcia minerálnych solí rastlinami, trans. s angličtinou, M., 1964; DA Sabinin, Vybrané práce o rastlinnej minerálnej výžive, M., 1971; Fyziológia koreňa, M., 1973.
D. B. Vakhmistrov.
Veľká sovietska encyklopédia M: "Soviet Encyclopedia", 1969-1978
Viete, že rastlina s koreňovým systémom absorbuje z pôdnych roztokov a potom asimiluje minerálne látky, ktoré potrebuje. Pre všetky rastliny je nevyhnutne potrebných 13 prvkov, ktoré sa podieľajú na metabolizme: dusík, fosfor, draslík, vápnik, horčík, síra, železo, mangán, meď, zinok, bór, molybdén, chlór. S úplnou absenciou jedného z týchto prvkov v pôde dochádza k prudkým zmenám v základných životných funkciách rastlín - rast je inhibovaný, normálny priebeh fotosyntézy je narušený. Dusík, fosfor, draslík, vápnik, síra a horčík sa nachádzajú v rastlinách vo významných množstvách a nazývajú sa makroelementy (z gréckeho slova „makro“ je veľké), zvyšok sú v zanedbateľných množstvách a nazývajú sa mikroelementy (z gréckeho slova „mikro“ je malé), ale sú pre závod veľmi dôležité.
Mnohé rastliny potrebujú iné prvky, ale nie sú pre nich absolútne nevyhnutné. Nevýznamné množstvá kobaltu zvyšujú rast sóje, zvyšujú obsah chlorofylu v listoch a vitamínu B i2 v uzlinách, ktoré sa tvoria na koreňoch rastliny, čo je veľmi dôležité pre aktivitu uzlových baktérií, ktoré obohacujú pôdu dusíkom.
Vysoká úroveň fotosyntézy závisí od správnej minerálnej výživy rastlín. Každý prvok súčasne plní špecifickú fyziologickú úlohu.
Napríklad dusík je priamo zapojený do tvorby prístroja, v ktorom dochádza k fotosyntéze; s účasťou fosforu v procese fotosyntézy sa tvoria organofosforové zlúčeniny bohaté na energiu; Draslík ovplyvňuje intenzitu fotosyntézy, akumuláciu chlorofylu a syntézu proteínov prebieha rýchlejšie.
Ak chcete poznať fyziologickú úlohu prvkov potrebných pre rastlinu, mechanizmus ich absorpcie a pohybu v rastline je veľmi dôležitý na správne aplikovanie hnojív.
Pestovateľ alebo pestovateľ plodín musí byť schopný včas zaznamenať nedostatok jedného alebo iného prvku v pôde. Dostupnosť pôdnych minerálnych prvkov môže byť určená chemickou analýzou pôdy, ako aj samotnými rastlinami. Na tento účel existuje mnoho spôsobov, ktorými sa môžete zoznámiť s tým, s čím sa môžete stretnúť v poľnohospodárskom podniku na farme, v štátnom statku alebo v inom špecialistovi. Nedostatok živín (alebo ich nadbytok) je možné vidieť jednoduchým okom zmenou farby listov, ich tvaru, prudkého spomalenia rastu a vývoja rastlín, pokiaľ, samozrejme, nie sú spôsobené inými príčinami: škodcami, chorobami, suchom, chladom.
Kyslá reakcia pôdy má tiež negatívny vplyv na rast a vývoj mnohých plodín.
Hlavné znaky nedostatkov výživy. S nedostatkom dusíka v rastlinách - bledozelená farba a žltnutie listov, slabý rast a skorý pokles listov; obilniny - slabé odnožovanie.
Ak je málo fosforu-tmavo-zelená alebo modrastá farba listu; objavia sa červené, fialové odtiene; tmavej, niekedy takmer čiernej farby sušiacich listov. S nedostatkom draslíka - žltnutie alebo zhnednutie listov, odumretie tkaniva alebo skrútenie po okrajoch listu, vrásky listov. Nedostatok síry je indikovaný svetle zelenou farbou listov bez uhynutia tkaniva. Nedostatok horčíka spôsobuje zosvetľovanie listov v dôsledku nedostatočnej tvorby chlorofylu, ako aj zmeny farby listov zo zelenej na žltú, červenú a fialovú na okrajoch a medzi žilami.
Nedostatok vápnika vedie k poškodeniu a smrti apikálnych pukov a koreňov.
Ak nie je dostatok železa, objaví sa jednotná žltkastosť medzi žilami listu alebo bledozelená a žltá farba listov bez odumretia tkaniva.
S nedostatkom bórových apikálnych pukov, koreňov a listov vymizne, nie je kvitnutie. Nedostatok medi spôsobuje zožltnutie listov, bielenie ich špičiek, prázdne zrno uší.
Niektoré rastliny, ktoré sú veľmi citlivé na nedostatok minerálnych prvkov, môžu slúžiť ako ukazovatele, či pôda je chudobná alebo bohatá na určité živiny. Nedostatok dusíka bude teda indikovaný predovšetkým karfiolom, brokolicou, bielou kapustou, jablkom, slivkou, čiernym ríbezlom; fosfor-repa, rutabaga, angrešt, jabloň; draslík - zemiaky, karfiol, fazuľa, fazuľa, egreše, ríbezle, jablone; horčík - karfiol, listová kapusta, zemiaky, egreše, čierne ríbezle, jablone; sodná repa, kŕmna repa; železo - rovnaké kultúry ako s nedostatkom dusíka a navyše ovos, zemiaky, jablká, hrušky, slivky, maliny; bórová cukrovka, krmivo, stôl, jablko; mangán-cukrová repa, krmivo, stôl, ovos, zemiaky, jablko, čerešňa, malina.
S cieľom kompenzovať nedostatok určitých látok v pôde sa do nej zavádzajú vhodné hnojivá, vrátane špeciálnych mikronutrientov obsahujúcich mikronutrienty, vo forme vrchných obväzov.
Je veľmi dôležité zaznamenať všetky prípady výskytu príznakov hladovania rastliny a vybrať oblasť na mape plodín, kde sú pozorované. Potom bude možné v tom istom roku nielen kŕmiť plodiny hnojivami, ale aj prijať opatrenia do budúcnosti na zlepšenie týchto pôd.
1. História vývoja teórie minerálnej výživy rastlín.
Prvé diela sa objavujú v stredoveku. Medzi nimi nazývame „Vedecký spis o rôznych pôdach (soli) a poľnohospodárstve“ francúzskeho prírodovedca Palissy, V tejto práci sa pôda po prvýkrát považuje za zdroj rastlinnej výživy s minerálnymi soľami, preto sa odporúča, aby sa minerálne látky vracali do pôdy vo forme hnojív.
Van gelmont, Experimentovali s rastlinami, na základe ktorých sa urobil chybný záver o kŕmení rastlín vodou. Willow a pot na zem (teória vody bola nesprávna)
Glauber predložila hypotézu, podľa ktorej základom rastu rastlín je dusičnan ("soľ plodnosti"), získaný zo zmesi hnoja a pôdy. Glauber navrhol, aby sa vytvoril z potravy zvierat. Odporúčal, aby sa pod vinicami vytváral liadok, zvlhčoval zasiate zrno roztokom liadku.
Lavoisier stanovil zákon o ochrane hmoty, určil zloženie vzduchu a proces tvorby oxidu uhličitého, urobil rad ďalších dôležitých objavov. Zároveň sa zaoberal aj otázkami súvisiacimi s agronomickou chémiou. Napísal: „Rastie materiál potrebný na ich organizáciu zo vzduchu, ktorý ich obklopuje, z vody a všeobecne z minerálneho kráľovstva“.
Priestley urobil nádherný zážitok. Vzal sklenenú čiapku, izoloval myš a horiacu sviečku pod ňou, potom dal mincovňu. Záver: Pod vplyvom zelených rastlín sa vzduch stáva vhodným pre dýchanie a pálenie.
Ingengaus zistil, že len rastliny, a len vo svetle, absorbujú oxid uhličitý zo vzduchu, pričom vyžarujú kyslík. Samotné rastliny nepretržite dýchajú, ale vo svetle vyžarujú oveľa viac kyslíka a pri neprítomnosti svetla používajú niektoré O2.
Senebier a Saussure prezentovali experimentálne dôkazy o minerálnej koreňovej a vzduchovej výžive rastlín. Vedci ukázali, že zelený škrob, tj chlorofyl, sa podieľa na absorpcii oxidu uhličitého. Vedci ukázali, že pod vplyvom slnečného žiarenia listy odparujú vlhkosť, čím uľahčujú prístup nových porcií „džúsov“ z pôdy a koreňov do nadzemných orgánov rastlín. Pri pôsobení svetla absorbujú oxid uhličitý a uvoľňujú kyslík. Korene pôdy dávajú rastlinám len malú časť potravy, ale tieto minerálne látky absolútne potrebujú.
Thayer bol zástancom humusovej teórie výživy rastlín. Podľa tejto teórie je humus priamo absorbovaný koreňmi a slúži ako hlavná potrava pre rastliny. Minerálne látky, táto teória bola mylne považovaná len za pomocnú, ktorá prispieva k absorpcii humusu.
Liebig ukázali, že všetky minerálne zlúčeniny rastliny sú absorbované z pôdy. Preto sa zotaviť úrodnosť pôdy tieto látky sa musia vrátiť do pôdy, „zákon návratu“; „Zákon minima“: úroveň úrody závisí od množstva minerálnej látky, ktorá je minimálna. Zvýšenie obsahu iných minerálnych látok zároveň nezvyšuje výťažnosť.
Boussingault dal niekoľko presných experimentov, ktoré dokázali, že rastliny nepoužívajú dusík obsiahnutý vo vzduchu. Po zistení, že ďatelina a lucerna obohacujú pôdu dusíkom, Boussingo predpokladal, že tieto rastliny absorbujú dusík zo vzduchu.
Popolné prvky sa koncentrujú v tých orgánoch a bunkách, ktorých úroveň vitálnej aktivity je pomerne vysoká. Pravidlom je, že čím bohatšia je pôda a suchšie podnebie, tým vyšší je obsah popola rastliny. Požadované položky. To sú tí, bez ktorých telo nemôže dokončiť svoj životný cyklus. Y. Sachs a I. Knoop zistili, že nasledujúcich 7 prvkov je nevyhnutných pre životne dôležitú aktivitu rastliny, okrem C, O, H, N, P, S, K, Ca, Mg, Fe. Neskôr sa ukázalo, že je potrebné, aby boli rastliny v šiestich ďalších prvkoch: bór, mangán, zinok, meď, molybdén a chlór. Pre niektoré rastliny sú charakteristické aj iné prvky (napríklad sodík). Všetky potrebné prvky koreňovej výživy podliehajú základným pravidlám Liebig. Všetky prvky potrebné pre životnosť rastlín, v závislosti od ich kvantitatívneho obsahu v rastline, sa zvyčajne delia na makronutrienty (obsah viac ako 0,01%) - N, P, S, K, Ca, Mg, Fe a stopové prvky (obsah menší ako 0,01%). Mn, C, Zn, B, Mo, O
Spoločné hodnoty živín: 1) sú zahrnuté v zložení biologicky dôležitých organických látok; 2) podieľať sa na tvorbe určitej iónovej koncentrácie, stabilizácii makromolekúl a koloidných častíc (elektrochemická úloha); 3) zúčastňujú sa katalytických reakcií, vstupujú do kompozície alebo aktivujú jednotlivé enzýmy.
3. Makroelementy - K, Ca, Mg, S, P, ich fyziologická úloha.
síra, Síra je absorbovaná rastlinami iba v oxidovanej forme, vo forme aniónového SCV. V rastline sa väčšina sulfátového aniónu redukuje na skupiny -SH a -S-S. Zahrnuté v troch aminokyselinách - cystín, cysteín a metionín, vitamíny, koenzýmy (biotín, tiamín, koenzým A, glutatión, kyselina lipoová). Podieľa sa na metabolizme proteínov, aeróbnej fáze dýchania, syntéze tukov, na tvorbe makroergickej väzby, metabolizme, atď.
Časť síry je v bunkovej šťave ako sulfátový ión. Keď koreň zomrie za podmienok, keď mu chýba kyslík, zlúčeniny obsahujúce síru sa rozkladajú na sírovodík, ktorý je jedovatý pre koreň. S nedostatkom síry, ako aj s nedostatkom dusíka začína ničenie chlorofylu, ale horné listy sú prvé, ktoré zažívajú nedostatok síry.
fosfor, Fosfor je absorbovaný rastlinami v oxidovanej forme vo forme solí kyseliny fosforečnej. Zahrnuté do nukleových kyselín (DNA a RNA), nukleotidov (ATP, NAD, NADP), nukleoproteínov, vitamínov, fosfolipidov (membrán) atď. Podieľa sa na procesoch glykolýzy a aeróbneho dýchania. Energia uvoľnená v týchto procesoch sa akumuluje vo forme fosfátových väzieb bohatých na energiu. Zúčastňuje sa fotosyntézy. V neprítomnosti fosforu na začiatku života a počas následného kŕmenia rastliny soľami fosforu, listy rastlín trpia určitý čas v dôsledku zvýšeného príjmu fosforu a metabolizmu dusíka v tejto súvislosti rušeného.
Draslík, vápnik a horčík absorbované z rozpustných solí, aniónov, ktoré nemajú toxický účinok alebo sú spojené s niektorou nerozpustnou látkou s kyslými vlastnosťami. Pre výživu sú nevyhnutné (K, Ca, Mg).
role: adsorbované na povrchu koloidných častíc protoplazmy, vytvárajú okolo nich určité elektrostatické sily, určujú obsah vody a vodnú silu koloidov.
draslík, Draslík v rastlinách je obsiahnutý vo veľkých množstvách, najmä v ich vegetatívnych častiach. Draslík má veľký vplyv na plazmatické koloidy, zvyšuje ich hydrofilitu (riedi plazmu). Draslík je tiež katalyzátorom mnohých syntetických procesov: spravidla katalyzuje syntézu vysokomolekulových látok z jednoduchších látok, podporuje syntézu sacharózy, škrobu, tukov, proteínov, ak je nedostatok draslíka, v procese sa narúšajú procesy syntézy a glukóza, aminokyseliny a produkty rozkladu iných vysokomolekulových zlúčenín. , S nedostatkom draslíka sa okrajová poistka objavuje na spodných listoch - hrany listovej čepele vymiznú, listy získajú charakteristický kopulovitý tvar a na listoch sa objaví nekróza.
vápnik, Niektoré z vápnika sú v bunkovej miazge. Tento vápnik sa aktívne nepodieľa na metabolických procesoch, zabezpečuje neutralizáciu nadbytku organických kyselín. Časť vápnika sa koncentruje v plazme, tu vápnik znižuje hydrofilitu plazmatických koloidov, zvyšuje ich viskozitu. Vápnik je súčasťou jadrovej hmoty, hrá úlohu v procesoch bunkového delenia. Podieľa sa na tvorbe bunkových membrán, tvorbe stien koreňových chĺpkov. V neprítomnosti vápnika sa rýchlo ovplyvňujú rastové body leteckých častí a koreňa, pretože vápnik sa nepohybuje zo starých častí rastliny na mladých. Korene sa priblížia, ich rast sa takmer zastaví alebo ide abnormálne.
magnézium, Zahrnuté v zložení chlorofylu. Horčík je dôležitý pre metabolizmus dýchacích ciest, katalyzuje rad reakcií tvorby fosfátových väzieb a ich prenosu. Pretože fosfátové väzby bohaté na energiu sa podieľajú na širokej škále syntéz, tieto procesy sa neuskutočňujú bez horčíka. Pri nedostatku horčíka sa molekula chlorofylu zničí a žilky listov zostanú zelené a oblasti tkanív medzi žilami sa zblednú. Tento jav sa nazýva flekatá chloróza.
železo, Absorbované vo forme rozpustených solí, komplexných a organických zlúčenín. V rastlinných tkanivách sa železo čiastočne premieňa na organické zlúčeniny. Keďže je súčasťou enzýmov, aktívne sa zúčastňuje na redoxných procesoch. Železo je súčasťou respiračných enzýmov (cytochróm, cytochróm oxidáza, kataláza a peroxidáza). Podieľa sa na tvorbe chlorofylu. S nedostatkom železa chloróza sa vyvíja. Pri nedostatku železa mladé listy menia farbu (žltnú), fotosyntézu; Rast rastlín sa spomaľuje.
4. Fyziologická úloha dusíka v rastlinnom živote. Zdroje dusíkovej výživy vyšších rastlín. Molekulárna fixácia dusíka.
Dusík je rastlinou dobre absorbovaný zo solí kyseliny dusičnej a amónia. Je hlavným prvkom výživy koreňov ako súčasť proteínov všetkých živých buniek. Dusík je integrálnou súčasťou nukleových kyselín, ktoré sú súčasťou jadra a sú nositeľmi dedičnosti. Je súčasťou chlorofylu, je súčasťou enzýmov, ktoré regulujú metabolické reakcie a množstvo vitamínov. Všetky formy dusíka v zariadení sa premieňajú na amoniakové zlúčeniny, ktoré pri reakcii s organickými kyselinami tvoria aminokyseliny a amidy - asparagín a glutamín. Amoniakálny dusík sa zvyčajne v zariadení vo významných množstvách nehromadí. Toto sa pozoruje len pri nedostatku sacharidov; Za týchto podmienok ho závod nemôže spracovať na neškodné organické látky - asparagín a glutamín. Nadbytok amoniaku v tkanivách často vedie k poškodeniu. Ak rastlina nemá dusík, potom to ovplyvňuje predovšetkým rýchlosť rastu. Nové výhonky sa takmer netvoria, veľkosť listov sa znižuje. V neprítomnosti dusíka v starých listoch je chlorofyl zničený. Biologická fixácia atmosférického dusíka je dôležitá, vďaka čomu dusík prechádza do foriem, ktoré môžu využívať všetky rastlinné organizmy a prostredníctvom nich živočíšne organizmy, organizmy schopné asimilovať dusík vo vzduchu je možné rozdeliť do skupín: 1) symbiotické dusíkaté fixátory - mikroorganizmy, ktoré sa asimilujú atmosférický dusík, ktorý je v symbióze s vyššou rastlinou; 2) nes symbiotické látky na fixáciu dusíka - mikroorganizmy, ktoré žijú voľne v pôde a absorbujú dusík zo vzduchu; 3) asociatívne dusíkové fixátory - mikroorganizmy žijúce na povrchu koreňového systému obilnín, t.j. žijúce v spojení s vyššími rastlinami. Dôležité sú symbiotické dusíkaté fixátory, ktoré žijú v uzlinách koreňov strukovín (baktérie uzlín) patriacich do rodu Rhizobium. azotobacterr. Aby tieto mikroorganizmy uskutočňovali proces fixácie dusíka, je nevyhnutná prítomnosť molybdénu, železa a vápnika. Konečným produktom fixácie dusíka je amoniak. V procese redukcie dusíka na amoniak je zahrnutý multienzýmový komplex nazývaný dusík. Zdrojom protónov a elektrónov na redukciu dusíka je transportný reťazec dýchacích elektrónov. To poukazuje na spojenie asimilácie atmosférického dusíka s procesmi dýchania, ako aj fotosyntézy (zdroj sacharidov). Na redukciu N2 na NH3 sa vyžaduje šesť elektrónov podľa rovnice: N2 + 6e + 2H + -\u003e 2NH 3 Tento proces vyžaduje ako zdroj energie ATP: 25-35 ATP molekúl sa spotrebuje, pretože dusíkáza je enzým s nízkou substrátovou špecifickosťou. Zvláštnosťou dusíkatej fázy je, že anaeróbne podmienky sú potrebné pre fungovanie enzýmu. Avšak v bunkách vyššej rastliny je na udržanie dýchania potrebný kyslík. Úlohou leggemoglobínu je viazať baktérie v tele a vytvárať podmienky pre fungovanie dusíkatej fázy. Fe, Cu a Co sú potrebné na tvorbu legemoglobínu. Pre normálny priebeh fixácie dusíka sú potrebné Mo a Fe, ktoré sú súčasťou dusíkovej fázy. Molybdén podporuje konformáciu dusíkatej bázy, podieľa sa na viazaní dusíka a prenosu elektrónov a tiež indukuje syntézu dusíkatej fázy. Kobalt je zložkou vitamínu B12, ktorý sa podieľa na procese biosyntézy legemoglobínu. Dusíkaté látky sa transportujú do nadzemných orgánov hostiteľskej rastliny hlavne vo forme amidov (asparagín, glutamín).
5. Metabolizmus dusíka vyšších rastlín: obnova dusičnanov a spôsob, akým asimilovať amoniak.
Vyššie rastliny absorbujú zlúčeniny dusíka z pôdy. Hlavným zdrojom výživy dusíka pre rastliny sú dusičnany a amoniak. Koreňové systémy rastlín sú dobre asimilované nitrátmi, ktoré vstupujú do koreňov rastliny, podliehajú enzymatickej redukcii na dusitany a ďalej na amoniak. Tento proces sa vyskytuje hlavne v koreňoch, ale tieto schopnosti majú aj listové bunky. Redukcia dusičnanov na amoniak prechádza radom krokov. V prvej fáze sa dusičnany redukujú na dusitany za účasti enzýmu nitrát reduktázy:
N03- + 2e -\u003e N02
Nitrát reduktáza je enzým obsahujúci FAD, hem a molybdén. Enzým je lokalizovaný v cytosóle, kde prebieha proces redukcie dusičnanov na dusitany. Dodávateľom NADH je proces respirácie a čiastočne aj svetelné reakcie fotosyntézy. Preto je obnova dusičnanov úzko spojená s výmenou dýchacích plynov a fotosyntézou. Na redukciu dusičnanov sa môžu použiť produkty tvorené v procese necyklickej fotofosforylácie (NADPH, ATP). Pri slabom osvetlení sa znižuje aktivita enzýmu Fe a Mo a v bunke sa hromadia nitráty.
Druhá fáza - redukcia dusičnanov na amoniak je katalyzovaná enzýmom nitrit reduktázou:
N02- + 6e -\u003e NH4 +
Nitrit reduktáza je enzým obsahujúci hém ako protetickú skupinu. Aktivita tohto enzýmu je vyššia ako aktivita dusičnanreduktázy. Nitrit reduktáza je lokalizovaná v listových chloroplastoch alebo koreňových plastidoch. Donorom elektrónov v listoch je redukovaný ferredoxín, ktorý vzniká pri fungovaní vo svetle PSI, a dusitany môžu vstupovať do rastliny z pôdy a podliehať redukcii na NH4. Pohyb dusitanov z cytoplazmy do chloroplastov je stimulovaný Ca. Tiež sa uskutočňuje reverzný proces - oxidácia amónnej formy dusíka na dusičnanovú formu, čo vyvracia rozsiahly názor na výhradne exogénny pôvod dusičnanov v rastlinách.
Najdôležitejším zdrojom výživy dusíka je amoniakálny dusík. Akumulácia amoniaku v bunkách vedie k nežiaducim následkom. Avšak rastliny majú schopnosť neutralizovať amoniak jeho pripojením k organickým kyselinám za vzniku amidov (glutamín, asparagín). Rastliny môžu byť rozdelené na amid, tvoriace amidy - asparagín a glutamín, a amóniové, tvoriace amónne soli. V procese respirácie sa ako medziprodukty tvoria organické kyseliny, vrátane a-ketoglutárovej a oxaloctovej. Tieto kyseliny ako výsledok priamej redukčnej aminačnej reakcie pridávajú amoniak:  Reakcia prebieha v dvoch stupňoch. V prechodnom stupni sa vytvorí aminokyselina. Reakcia je katalyzovaná enzýmom glutamát dehydrogenázou s aktívnou skupinou NAD. Tento enzým je lokalizovaný hlavne v mitochondriách, ale môže byť obsiahnutý v cytosóle a chloroplastoch. Kyselina asparágová sa vytvára analogicky s kyselinou glutámovou redukčnou amináciou kyseliny oxaloctovej za účasti enzýmu aspartátdehydrogenázy. Aktivita glutamínsyntetázy je ovplyvnená prítomnosťou katiónov: Mg2 +, Mn2 +, Co2 +, Ca2 +. Enzým sa nachádza v cytosóle, ale možno ho nájsť aj v chloroplastoch. Asparagín sa vyrába podobným spôsobom. V mladších orgánoch (listoch) a dokonca aj v mladších bunkách je tvorba amidov intenzívnejšia. Úloha amidov v rastlinách je rôzna. Nie je to len forma neutralizácie amoniaku, je to tiež transportná forma dusíkatých zlúčenín, ktorá zabezpečuje ich odtok z jedného orgánu do druhého. Je dôležité, aby amidy a ich bezprostredné prekurzory - kyselina glutámová a kyselina asparágová - boli materiálom na budovanie mnohých ďalších aminokyselín v procesoch transaminácie, ako aj preusporiadanie ich uhlíkovej štruktúry.
Reakcia prebieha v dvoch stupňoch. V prechodnom stupni sa vytvorí aminokyselina. Reakcia je katalyzovaná enzýmom glutamát dehydrogenázou s aktívnou skupinou NAD. Tento enzým je lokalizovaný hlavne v mitochondriách, ale môže byť obsiahnutý v cytosóle a chloroplastoch. Kyselina asparágová sa vytvára analogicky s kyselinou glutámovou redukčnou amináciou kyseliny oxaloctovej za účasti enzýmu aspartátdehydrogenázy. Aktivita glutamínsyntetázy je ovplyvnená prítomnosťou katiónov: Mg2 +, Mn2 +, Co2 +, Ca2 +. Enzým sa nachádza v cytosóle, ale možno ho nájsť aj v chloroplastoch. Asparagín sa vyrába podobným spôsobom. V mladších orgánoch (listoch) a dokonca aj v mladších bunkách je tvorba amidov intenzívnejšia. Úloha amidov v rastlinách je rôzna. Nie je to len forma neutralizácie amoniaku, je to tiež transportná forma dusíkatých zlúčenín, ktorá zabezpečuje ich odtok z jedného orgánu do druhého. Je dôležité, aby amidy a ich bezprostredné prekurzory - kyselina glutámová a kyselina asparágová - boli materiálom na budovanie mnohých ďalších aminokyselín v procesoch transaminácie, ako aj preusporiadanie ich uhlíkovej štruktúry.
6. Stopové prvky v rastlinnom živote.
Okrem základných prvkov je pre rast rastlín potrebný celý rad tzv. Mikroelementov. Stopové prvky sa absorbujú len pri nízkych koncentráciách zodpovedajúcich solí. S rastúcimi dávkami sú už jedovaté pre rastlinu. Ich úloha v živote rastlín, ako sú vitamíny, je spojená s aktivitou enzýmov.
bor, Aby sa zariadenie mohlo normálne vyvíjať, musí byť zásobované bórom. V neprítomnosti bóru sa zastaví rast koreňov a spodná časť. Rastové body odumierajú, ako sa bunky mladého rastúceho tkaniva - meristémy prestávajú deliť. Bór sa zúčastňuje procesu klíčenia peľu a rastu vaječníkov, preto sa s jeho nedostatkom prudko znižuje produkcia semien rastlín a hrá veľkú úlohu v pohybe cukrov; Mnohé zlúčeniny organického bóru sú rastové stimulátory.
Meď. Veľká časť medi sa koncentruje na chloroplasty. Zdá sa, že meď katalyzuje niektoré reakcie vo fotosyntéze, s nedostatkom medi sa ukáže, že chloroplast je krátkodobý. Meď je súčasťou mnohých oxidačných enzýmov (polyfenol oxidáza, tyrozináza, atď.) Meď hrá významnú úlohu v metabolizme proteínov.
zinok, Zinok je súčasťou dôležitého enzýmu - karbohydrát. Okrem toho sa zinok zúčastňuje syntézy aminokyseliny tryptofánu, ktorá je prekurzorom auxínov v rastline.
mangán, Katalyzuje arbilylačné reakcie a hrá dôležitú úlohu pri fotosyntéze a dýchaní. Väčšinou sa akumuluje v listoch av miestach rastu. Prítomnosť mangánu v živnom roztoku prispieva k oxidačným transformáciám a zvyšuje sa tak vstrebávanie dusičnanového dusíka. Zvlášť charakteristickým znakom mangánu je jeho schopnosť oxidovať zlúčeniny železa.
molybdén, Molybdén je nevyhnutný pre rastliny v extrémne malých množstvách. Katalyzuje procesy redukcie dusičnanov a syntézy proteínov.
7. Mechanizmus vstupu iónov do bunky. Úloha bunkovej steny. Iónový transport cez membrány: pasívny a aktívny.
Absorpcia iónov bunkou začína ich interakciou s bunkovou stenou. Ióny môžu byť čiastočne lokalizované v intermikulárnych a interibrilárnych medzerách bunkovej steny, fixovaných v bunkovej stene elektrickými nábojmi. Objem buniek dostupný pre voľnú difúziu iónov sa nazýva voľný priestor. Voľný priestor zahŕňa medzibunkové priestory, bunkové steny a medzery, ktoré sa môžu vyskytnúť medzi bunkovou stenou a plazmatickou membránou. Niekedy sa to nazýva zdanlivý voľný priestor (PCB). Výraz „zdanlivý“ znamená, že objem závisí od predmetu a povahy rozpustenej látky. Voľný priestor rastliny sa nazýva apoplast, simplast - súbor protoplastov všetkých buniek. Absorpcia a vylučovanie látok v PSC je fyzikálno-chemický pasívny proces, ktorý nie je závislý od teploty a inhibítorov metabolizmu energie a proteínov. Bunková stena má vlastnosti iónomeniča, pretože v nej sú adsorbované ióny H + a HCO - 3, ktoré sa vymieňajú v ekvivalentných množstvách za ióny externého roztoku. Aby sa prenikla do cytoplazmy a zapojila sa do metabolizmu, ióny musia prejsť cez plazmatickú membránu. Iónový transport cez membránu môže byť pasívny a aktívny. Pasívna absorpcia nevyžaduje energiu a uskutočňuje sa difúziou pozdĺž koncentračného gradientu látky, pre ktorú je plazmatická membrána priepustná. Pasívny pohyb iónov je určený nielen chemickým potenciálom m, ako je to pri difúzii nenabitých častíc, ale aj elektrickým potenciálom e. Oba potenciály sú spojené vo forme elektrochemického potenciálu `m:` m = m + nFe, kde m je chemická látka, e je elektrická, `m je elektrochemický potenciál, n je valencia iónu, F je Faradayova konštanta. Potrebujete to?
Elektrický potenciál na membráne - transmembránový potenciál sa môže vyskytnúť z nasledujúcich dôvodov: 1) ak iónový prívod sleduje koncentračný gradient, ale vzhľadom na rozdielnu permeabilitu membránových katiónov, ako anióny, dochádza k vyššej rýchlosti. Z tohto dôvodu vzniká na membráne rozdiel v elektrických potenciáloch, čo vedie k difúzii opačne nabitého iónu; 2) ak sú proteíny na vnútornej strane membrány, ktoré fixujú určité ióny; 3) ako výsledok aktívneho transportu buď katiónu alebo aniónu, v tomto prípade sa opačne nabitý ión môže pohybovať pasívne pozdĺž gradientu elektrického potenciálu.
Aktívny transport je transport, ktorý ide proti elektrochemickému gradientu s výdajom energie uvoľnenej v procese metabolizmu. Ako sa teplota zvyšuje, jedy inhibujú. Zvýšenie ATP zvyšuje absorpčný proces. Aktívny transport iónov cez membránu sa uskutočňuje pomocou nosičov. Ión reaguje s nosičom na povrchu plazmatickej membrány. Nosičový komplex s iónom je mobilný v samotnej membráne a pohybuje sa na svoju vnútornú stranu. Tu sa komplex rozkladá a ión sa uvoľňuje do vnútorného prostredia a nosič sa presúva na vonkajšiu stranu membrány. Doprava zahŕňajúca nosiče môže sledovať gradient elektrochemického potenciálu. Ide o pasívny transport, ale vďaka nosičom ide s väčšou rýchlosťou ako obyčajná difúzia a tento proces sa nazýva rozptyl svetla. Na použitie energie ATP sa musí hydrolyzovať: ATP + HON ® ADP + Fn. Katalyzované pomocou ATPázy. Transportné ATPázy sú lipoproteíny s vysokou molekulovou hmotnosťou. Pre rastliny má veľký význam H + - ATPáza (vodíkové čerpadlo alebo vodíkové čerpadlo), prenos vodíkových iónov je sprevádzaný prenosom katiónov v opačnom smere, ktorý sa nazýva antiport, ktorý sa môže pohybovať spolu s protónom. ATPáza, pričom enzým, ktorý viaže špecifický ión, sa otáča a otáča na druhej strane membrány.
Intracelulárny transport iónov sa uskutočňuje v dôsledku pohybu cytoplazmy a kanálov endoplazmatického retikula. Ióny vstupujú do vakuoly, ak sú s nimi cytoplazmy a organely už nasýtené. Aby sa ióny dostali do vakuoly, musia prekonať ďalšiu bariéru - tonoplast. Iónový transport cez tonoplast sa tiež vykonáva pomocou nosičov a vyžaduje energiu.
8. Koreň ako orgán absorpcie minerálnych prvkov. Metabolizmus koreňov.
Hlavnou zónou absorpcie živín, zásobujúcich a rastúcich orgánov, je zóna napínania buniek a zóna koreňových chĺpkov. V meristematickej zóne nie je žiadny diferencovaný vaskulárny systém. Súčasne je floém diferencovaný skôr a len xylem sa tvorí len mierne vyššie pozdĺž dĺžky koreňa. Je na xyléme, že voda je transportovaná rozpustenými živinami. Preto je v týchto bunkách použitý objem absorbovaných iónov meristému.
Nad zónou koreňových chĺpkov je rozvetvená zóna koreňa. V tejto zóne je povrch pokrytý vrstvou korku a prakticky sa nepodieľa na absorpcii výživných solí. Rôzne zóny koreňa absorbujú rôzne minerálne prvky. Ukázalo sa, že Ca2 + vstupuje iba do apikálnych zón, K +, NH4 + a fosfáty sú absorbované celým koreňovým systémom.
Vlastnosti prijímania: schopnosť rastlín selektívne koncentrovať látky; relatívna nezávislosť vody a soli; závislosť na dýchaní a fotosyntéze; urýchlenie procesu pod vplyvom teploty a svetla.
V koreňovom systéme sa rozlišujú dva zväzky - apoplast a symplast. Väčšina pôdnych koloidov je záporne nabitých, katióny sú na povrchu v adsorbovanom (absorbovanom) stave. Katióny a anióny, ktoré sú v absorbovanom stave na časticiach komplexu absorbujúceho pôdu, môžu byť vymenené za ióny adsorbované na povrchu koreňových buniek. Týmto spôsobom môžu byť K +, Ca2 +, Na + katióny dodávané výmenou za protóny, ako aj N03-, P043- a ďalšie anióny výmenou za HC03- alebo anióny organických kyselín. Absorbované ióny sú adsorbované na povrchu stien rhododermálnych buniek. Z adsorbovaného stavu sa môžu ióny pohybovať pozdĺž kortexu koreňa dvoma spôsobmi: pozdĺž apoplastu a symplastu. Po vstupe do symplastu prenikajú ióny cez membránu a potom sa pohybujú pozdĺž plazmidov do xylemových ciev.
Prúd vody so solutami pohybujúcimi sa vo voľnom priestore (apoplast), ako to boli všetky bunky kortexu. Počas tejto dráhy je možné pozorovať adsorpciu látok bunkovými stenami, vstup iónov do buniek cez vhodné membrány a ich začlenenie do metabolizmu, t.j. metabolizácie. Treba poznamenať, že v dráhe pohybu pozdĺž apoplastu vo forme endodermálnych buniek s pásmi Caspari je obštrukcia. Pohyb cez endodermálne bunky je možný iba prostredníctvom cytoplazmy. V tomto ohľade je nevyhnutný prenos iónov cez membrány endodermálnych buniek a tiež s pomocou nosičov. Vo všeobecnosti sa v koreňovom systéme medzi apoplastom a symplastom vyskytuje kontinuálna interakcia a výmena výživných solí.
9. Ekológia minerálnej výživy: vplyv vonkajších a vnútorných faktorov. Fyziologický základ aplikácie hnojiva.
Vplyv vonkajších podmienok
Pri teplote blízkej 0 ° C je absorpcia solí pomalá, potom až do 40 ° C sa zvyšuje. Zvýšenie teploty o 10 ° C môže spôsobiť zvýšenie absorpcie dvoch a dokonca trikrát. V tme sa absorpcia solí spomaľuje a postupne sa zastavuje a zrýchľuje pod vplyvom svetla. Zníženie koncentrácie kyslíka pod 3% spôsobuje pokles absorpcie približne dvakrát. V prítomnosti ľahko absorbovaného aniónu prichádzajú katióny rovnakej soli rýchlejšie. Ióny s rovnakým nábojom si zvyčajne konkurujú.
Vplyv vnútorných faktorov
Závisí od intenzity dýchania. Proces dýchania môže ovplyvniť tok solí v niekoľkých smeroch. V procese respirácie sa teda uvoľnený oxid uhličitý vo vodnom prostredí disociuje na ióny H + a HC03–. Tieto ióny sú adsorbované na povrchu koreňa a slúžia ako výmenný fond pre prichádzajúce katióny a anióny. Prenos iónov cez membránu zahŕňa špecifické nosné proteíny, ktorých syntéza závisí od intenzity respiračného procesu.
Racionálne zavádzanie živín vo forme hnojív je významným faktorom pri zvyšovaní výnosu rastlín. Treba však mať na pamäti, že nadmerné dávky hnojív sú pre ich odpad nielen zbytočné, ale môžu viesť k mnohým veľmi škodlivým následkom. Predovšetkým môže vytvoriť zvýšenú koncentráciu pôdneho roztoku. Väčšina pestovaných rastlín je na tento ukazovateľ citlivá. Zvýšenie obsahu akejkoľvek živnej soli môže mať priamy toxický účinok na telo rastliny. Ak chcete stanoviť primerané sadzby hnojív, je potrebné vziať do úvahy prítomnosť živín v pôde, potreby tejto rastliny, vlastnosti hnojív, zloženie vylučovania koreňov, transpiračných koeficientov, fotosyntéza, kyslosť pôdy.
MINERÁLNA SILA RASTLÍN MINERÁLNA SILA RASTLÍN
súbor procesov absorpcie, pohybu a asimilácie chem. prvky potrebné pre rast života, telo, vo forme iónov minerálnych solí. Medzi prvkami M. p. rozlišovať makronutrienty (N.S.P, K, Ca, Mg) a mikroelementy. Dusík je absorbovaný rastlinami vo forme aniónu NO3 alebo katiónu NH +, fosforu a síry vo forme aniónov H2PO4 a SO2-, kovov - vo forme katiónov K +, Ca2 + a Mg2 +. Jednobunkové organizmy a vodné rastliny absorbujú prvky M. pp. celý povrch, vyššie zemské rastliny - povrchové bunky koreňa, ch. ARR. koreňové chĺpky. Cementy prenikajú bunkou cez plazmatickú membránu, ktorá ju obklopuje, pasívne, anióny (rovnako ako K + pri vonkajších koncentráciách nižších ako 1 mM) sú aktívne, s metabolickými nákladmi. energie. Aktívna absorpcia je zabezpečená prácou molekulárnych iónových čerpadiel plazmatickej membrány. Vo vnútri bunky sa ióny zmiešavajú pomocou kruhového pohybu cytoplazmy (cyklozy) a sú medzi nimi a organoidmi (kompartment) nerovnomerne redistribuované. Od buniek k bunkám sa ióny pohybujú buď pozdĺž plazmidovej kaše, ktorá spája všetky bunky tkaniva dohromady - do symplastu, alebo pozdĺž bunkových membrán, tiež spojených do apoplastu. V tele rastliny sa ióny pohybujú prúdom vody cez xylémové cievy, ktoré prenikajú koreňom, stonkou a listom. Vzostupné dopravné prvky M. pp. nasmerovaný ch. ARR. vznikajúcich plodov a mladých listov. So starnutím starším. listové prvky M. pp z nich do rastúcich orgánov, kde ich možno opätovne použiť (recyklácia). Zapojenie absorbovaných prvkov M. pp. Vo všeobecnosti dochádza k metabolizmu vo všetkých rastlinných bunkách. Dusík je súčasťou aminokyselín a proteínov, ako aj amínov, amidov, alkaloidov, chlorofylu, nukleových kyselín, nukleotidov, pl. hormónov a vitamínov. Síra je zahrnutá v aminokyselinách cysteín, cistnn a metionín, fosfor - v adenozíntrifosfáte (ATP) a ďalších adenozín fosfátoch, ktoré hrajú kľúčovú úlohu v energetich. výmenu buniek a tiež vo fosfolipidoch bunkových membrán a v nukleových bunkách. Draslík, vápnik a horčík zostávajú Ch. ARR. v iónovej forme, zabezpečujúcej stabilitu subcelulárnych štruktúr a aktivitu enzýmov (približne 10% buniek Mg listov je súčasťou chlorofylu). Spolu s fotosyntézou M. p. robí jednotný proces výživy rastlín. Predpis M. p. Použitie hnojív je jedným z najdôležitejších spôsobov, ako zvýšiť produktivitu poľnohospodárskych produktov. kultúry.
.(Zdroj: "Biological Encyclopedic Dictionary." Editor-in-Chief, M. S. Gilyarov, Redkol: A. A. Babaev, G. G. Vinberg, G. A. Zavarzin a kol., 2. vyd., Corrected) - M .: Sov. Encyklopédia, 1986.)
minerálna výživa rastlínSúbor procesov absorpcie, pohybu a asimilácie chemických prvkov rastlinami, získaných z pôdy vo forme iónov minerálnych solí. V štúdii rastlinného popola sa v ňom našlo mnoho chemických prvkov, vrátane. zriedkavé, ktorých obsah v rôznych častiach rastlín nebol rovnaký. To znamená, že tieto prvky sú potrebné pre rastliny a akumulujú sa v nich. Prvky prítomné vo všetkých rastlinách boli považované za životne dôležité - jedná sa o draslík, vápnik, horčík, železo, síru a fosfor. Pre rôzne rastliny sú potrebné v rôznych množstvách. Nie je možné úplne nahradiť jeden prvok iným. Výnos poľnohospodárskej produkcie závisí od stupňa ich prítomnosti v pôde. rastliny. V pôde stredného Ruska zvyčajne nemajú dostatok dusíka a kyseliny fosforečnej, menej často draslíka, preto sa zavádzajú ako dusík a fosfor-draselné hnojivá.
Každý chemický prvok hrá osobitnú úlohu v živote rastliny. Fosfor je rastlinou absorbovaný vo forme solí kyseliny fosforečnej (fosforečnanov) a je v ňom vo voľnom stave alebo spolu s proteínmi a inými organickými látkami, ktoré sú súčasťou plazmy a jadra. Vo voľnom stave je možné regulovať kyslé a zásadité prostredie v bunke. Síra je rastlinou absorbovaná vo forme solí kyseliny sírovej, je súčasťou proteínov a éterických olejov. Draslík sa koncentruje v mladých orgánoch bohatých na plazmu, ako aj v orgánoch akumulácie rezervných látok - semená, hľuzy, pravdepodobne hrajú úlohu neutralizátora kyslej reakcie bunkovej miazgy a zúčastňujú sa turgoru. Horčík sa nachádza v rastline na rovnakom mieste ako draslík a navyše je súčasťou chlorofylu. Vápnik sa hromadí v dospelých orgánoch, najmä v listoch, slúži ako neutralizátor kyseliny šťaveľovej škodlivej pre rastlinu a chráni ju pred toxickými účinkami rôznych solí a podieľa sa na tvorbe mechanických membrán. Železo sa nachádza v rastline v malých množstvách, ale je súčasťou protoplastov, a s jeho nedostatkom sa vyvíjajúce listy nezmenia na zelené, ale zostávajú biele (fenomén chlorózy).
Okrem týchto životne dôležitých prvkov má chlorid sodný určitú hodnotu (akumuluje sa v bunkách halofilné Rastlinyumožňuje zvýšiť osmotický tlak na 100 atmosfér, takže môžu odolať fyziologickej suchosti pôdy), mangánu, fluóru, jódu, brómu, zinku, kobaltu, stimulácii rastu rastlín a ďalšie.
Minerálne zlúčeniny prvkov dusíka a popola sú absorbované suchozemskými vyššími rastlinami takmer výlučne koreňmi. Soli, podobne ako voda, sú absorbované živými bunkami primárneho kortexu koreňových a koreňových chĺpkov, potom sú tlačené koreňovým tlakom do nádob s vodou, transportované inými časťami rastliny a opäť odobraté živými bunkami stonky a listu. V živých bunkách koreňa sa objavuje prvá voľba látok povolených vo vnútri rastliny. Účasť živých buniek na prijímaní látok určuje selektívnu schopnosť rastliny, vďaka ktorej sa rôzne látky absorbujú v rôznych množstvách. Keďže dodávka je vysoko závislá na spotrebe, rastlina prijíma niektoré soli v iných štádiách vývoja, potom iné. Čím bližšie je kontakt koreňa s pôdnymi časticami, tým viac sa vytvoril koreňový systém a čím viac je absorpcia solí úplnejšia. Okrem toho majú korene schopnosť rozpúšťania. Silný, vysoko rozvetvený koreňový systém nepochybne prispieva k lepšej výžive rastlín.
.(Zdroj: "Biológia. Moderná ilustrovaná encyklopédia." Hlavný redaktor A. P. Gorkin; M.: Rosmen, 2006.)
minerálna výživa rastlín - Absorpcia a absorpcia živín rastlinami v minerálnej forme. [GOST 20432 83] Témy hnojív ... Technický prekladateľ
Z vonkajšieho prostredia asimilovať ióny minerálnych solí, ktoré sú nevyhnutné pre normálne fungovanie rastlinného organizmu. K prvkom M. p. Medzi príklady patria N, P, S, K, Ca, Mg, ako aj stopové prvky (Fe, B, Cu, Zn, Mn, atď.). M. p. P. ... ...
minerálna výživa rastlín - minerálna výživa rastlín, súbor procesov absorpcie, pohybu a asimilácie chemických prvkov rastlinami z pôdy vo forme iónov minerálnych solí. K prvkom M. p. zahŕňajú makro (N, S, P, K, Ca, Mg) a stopové prvky ... ...
MINERÁLNA SILA RASTLÍN - súbor procesov absorpcie, pohybu a asimilácie chemickej látky. prvky rastlín z pôdy vo forme ionoper miner, soli. K prvkom M. p. Makroelementy (N, S, P, K, Ca, Mg) a mikroprvky (Fe, B, Mn, Cu, Zn, Mo) sú zahrnuté. Dusík sa vstrebáva ... ...
Proces absorpcie a asimilácie rastlinami z prostredia chemických prvkov potrebných pre ich život; je presunutie látok z prostredia do cytoplazmy rastlinných buniek a ich chemická transformácia na zlúčeniny, ... ... Veľká sovietska encyklopédia
výživy rastlín - výživa rastlín, asimilácia anorganických zlúčenín z prostredia a ich autotrofná transformácia na organické látky používané na tvorbu a obnovu konštrukčných častí rastlín a na dodávku energie do funkcií. Až do začiatku ... Poľnohospodárstvo. Veľký encyklopédický slovník
NAPÁJANIE - Anorganická absorpcia. zlúčeniny z prostredia a ich autotrofná transformácia na orgán. vo VA, používané na vzdelávanie a aktualizáciu konštrukčných častí riešenia a energie. poskytovanie funkcií. Až do začiatku 19. storočie bola tu teória humusu ... ... Poľnohospodársky encyklopédický slovník
Asimilácia (asimilácia) živín rastlinami pochádzajúcimi z vonkajšieho prostredia; metabolizmu. Zdroje živín pre rastliny je pôda, z ktorej sa rozpúšťajú v minerálnej vode (pozri ... ... Biologický encyklopédický slovník
Spotreba rastlín a zvierat (zvyčajne trofejou) minerálnych látok (vo forme aniónov alebo katiónov) nevyhnutných pre ich životne dôležitú činnosť. Ekologický encyklopédický slovník. Kišiňov: Hlavná redakčná rada moldavskej ... ... Ekologický slovník
MINERÁLNE POTRAVINY - absorpcia a asimilácia (asimilácia) koreňmi rastlín anorganických zlúčenín makro a mikroelementov, ako aj ich pohyb a transformácia v nadzemných častiach rastlín ... Botanický slovník
Newchemistry: Aké transfúzne systémy vyrábate? JSC "Syntéza", Kurgan: Vyrábame systémy pre ...
Nevyhnutnou zručnosťou lekárskych špecialistov je odber krvi zo žily. A táto schopnosť musí byť sprevádzaná ...
Všetko, čo súvisí s operáciou a účinkom anestézie, sa zvyčajne označuje ako „operačný stres“ a jeho dôsledky ...
Transfúzne systémy Infúzne systémy s plastovými a kovovými ihlami: Infúzne systémy sú navrhnuté tak, aby ...
Liečba a prvá pomoc pri extravazácii - pod kožu. Rozsah intervencie závisí od štádia extravazácie, ...
[0001] Vynález sa týka medicínskej technológie a môže byť použitý v transfúzii na transfúziu krvi, jeho ... \\ t
Infúzna terapia je liečba založená na intravenóznom alebo subkutánnom podaní rôznych ...

Budete potrebovať - statív, - fľašu liekov, - ampulky s drogami, - injekčnú striekačku, - 70 stupňov ...
Existuje mnoho rôznych chorôb, v ktorých sa proces liečby aj proces ďalšieho oživenia ...

Strana 17 z 31 17 Použitie roztokov koloidov a kryštaloidov v resuscitácii ...

Skúsení záhradníci vedia, že paradajka je jemná kultúra a mimoriadne náročná na živiny, zatiaľ čo ...
Doma, dať kvapkanie bez riadnych skúseností a konzultácie s lekárom nemôže byť.
Ako používať kvapkadlo? Sú situácie, keď pes alebo mačka potrebuje intravenózne infúzie, je to jednoduchšie ...
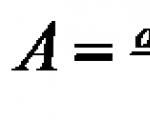
Hnojenie je veda. Zároveň je potrebné poznať také funkcie pôdy ako jej schopnosť vyrovnávať ...
Jedna z hlavných patogenéz väčšiny kritických stavov (akútna strata krvi, sepsa, šok ...
Vážení kolegovia, v tomto článku by som rád vyjadril základné princípy vedenia infúznej terapie (IT) ...